固有免疫记忆——训练免疫的研究进展①
2019-05-27李京蔓潘宇晨侯亚义
李京蔓 潘宇晨 窦 环 侯亚义
(南京大学生命科学学院,南京210093)
1 背景介绍
经典的免疫系统分为固有免疫和获得性免疫。固有免疫系统中效应细胞(如单核细胞、巨噬细胞、中性粒细胞和NK细胞)介导的免疫反应是迅速、非特异性和非记忆性的,而获得性免疫的建立是慢速、抗原特异性且具有免疫记忆的。以往认为只有获得性免疫具有免疫记忆,然而近年来有研究表明,在缺乏适应性免疫的植物和无脊椎动物以及某些哺乳动物中,机体在面对二次感染时也会表现出更强的抵抗力[1],不同病原体之间还会产生交叉保护作用[2],这种现象被称为“训练免疫”或“固有免疫记忆”,即某些刺激能够持久地激活先天免疫细胞,尤其是髓系细胞,诱导它们对未来的刺激做出更强烈的应答,但不具备特异性[3]。本文将具体介绍训练免疫的表现、机制以及在疾病中的重要作用。
1.1训练免疫概况 自2011年“训练免疫”这一概念提出后[4],越来越多的研究都证实了训练免疫的存在。人类单核细胞在体外接触微生物的组成成分如白色念珠菌细胞壁上的β-葡聚糖或卡介苗进行“训练”,一周后再次给予其他刺激,这些“经过训练”的细胞分泌细胞因子的能力会高于未经训练的细胞[5,6]。此外,给严重联合免疫缺陷(Severe combined immunodeficiency,SCID)小鼠体内注射卡介苗进行“训练” 后,致命白色念珠菌再感染所致的死亡率降低[7]。对人类来说,BCG能够对单核细胞进行训练,增强它们对微生物的反应性,这或许能在一定程度上解释接种疫苗的儿童受各种传染病致死的几率降低的现象[5,7-10]。
在经典的获得性免疫记忆当中,免疫系统会识别特异性抗原,T淋巴细胞和B淋巴细胞通过基因重组创造出众多具有精细识别抗原能力的抗原受体,即B细胞特异性抗原识别受体(B cell receptor,BCR)和T细胞特异性抗原识别受体(T cell receptor,TCR),这些淋巴细胞发生特异性扩增,清除异物,随后产生记忆细胞,对下一次相同外来抗原的刺激快速做出反应[11],相比于第一次反应,获得性免疫记忆所引发的反应特异性更强、强度更大。在上述对单核细胞“训练”的研究中,人们发现训练免疫和获得性免疫记忆有很大区别。首先,训练免疫涉及的细胞、识别受体和效应分子与经典免疫记忆不同。其次,训练免疫应对二次刺激时,虽然免疫反应性更强,但这种反应不具有特异性,而且受到转录因子和表观遗传学调控,即总体表现为转录过程和细胞生理机能的持续性变化,但并不会引起基因组发生基因突变和重组。最后,训练免疫细胞内的代谢过程会发生改变,某些代谢物的积累可以影响表观遗传,从而使细胞产生训练免疫表型[3](图1)。
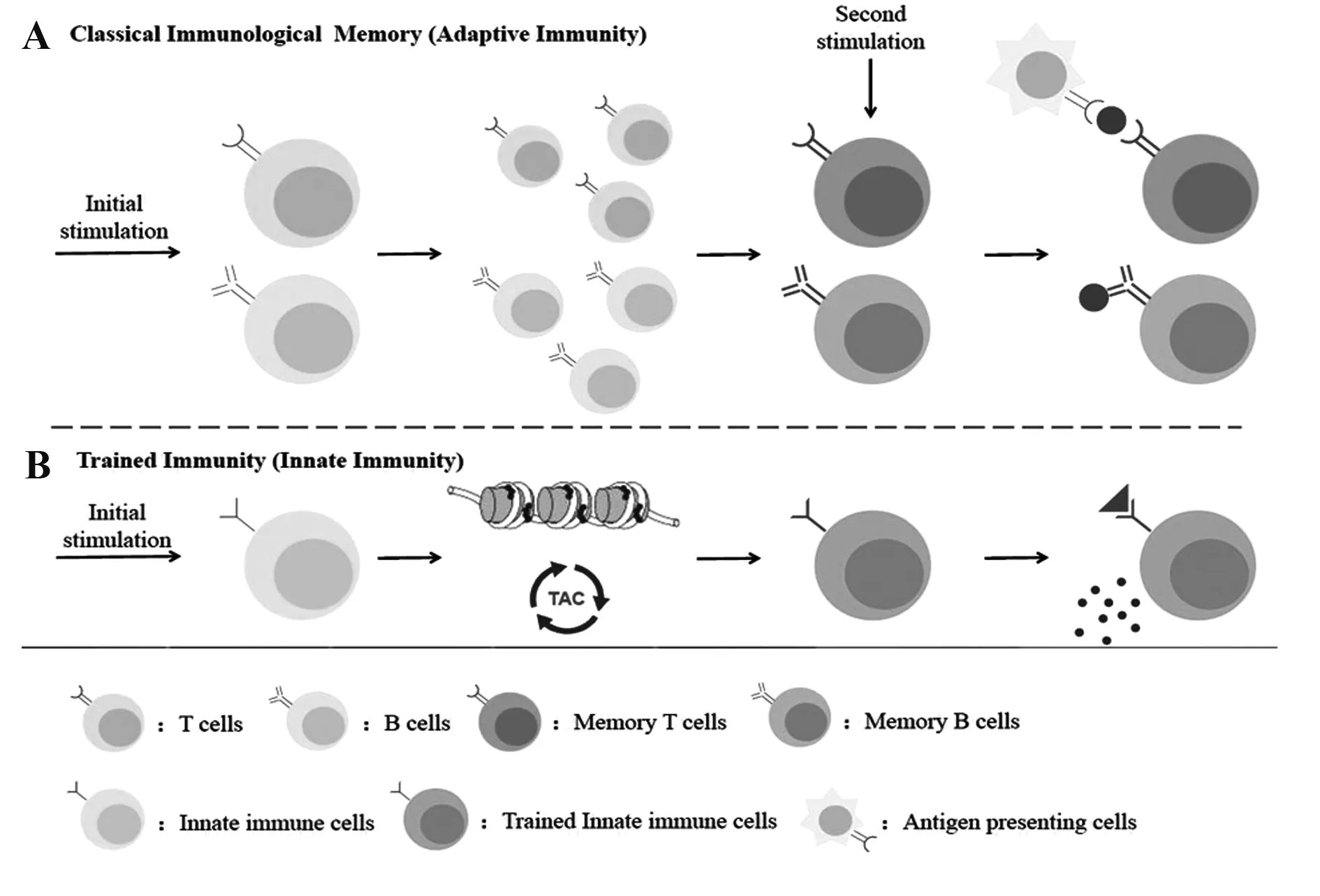
图1 经典的免疫记忆(获得性免疫)与训练免疫记忆(固有免疫)之间的比较Fig.1 Comparison between classical immunological memory (adaptive immunity) and trained immunity(innate immunity)
2 训练免疫相关机制
目前认为,训练免疫是免疫、代谢和表观遗传三者之间相互作用的结果。固有免疫细胞代谢的改变影响了表观遗传,后者又能进一步影响代谢通路和细胞因子的产生。下面我们将从细胞、表观遗传学和代谢途径三个方面对训练免疫的机制进行阐述。
2.1细胞种类 具有免疫记忆能力的固有免疫细胞有很多,主要包括单核细胞、巨噬细胞和自然杀伤细胞(Natural killer cells,NK cells)。与淋巴细胞不同,固有免疫细胞不会发生抗原受体基因的重排,而是通过表达模式识别受体(Pattern recognition receptors,PRRs)等,对外来抗原的结构——抗原相关分子模式(Rathogen-associated molecular patterns,PAMPs)和内源性危险信号——损伤相关分子模式(Damage-associated molecular patterns,DAMPs)进行识别和应答[12,13]。最初,人们从分子水平上研究巨噬细胞对细菌脂多糖(Lipopolysaccharide,LPS)的耐受现象,推断巨噬细胞可能具有记忆性[14]。后来,在免疫耐受方面研究较多的细胞如单核细胞、巨噬细胞和NK细胞上都发现了免疫记忆的现象,即细菌、真菌、寄生虫和病毒的感染均能触发单核细胞和巨噬细胞的训练免疫[15,16]。细胞因子(例如IL-12、IL-15和IL-18) 组合刺激可以诱导记忆性NK细胞的产生[17],这种NK细胞具有免疫记忆、能够长期存活并具有介导再次免疫应答的能力[18],从接种BCG的个体分离得到的NK细胞,用分枝杆菌和其他无关抗原刺激后,产生促炎因子水平增强[5]。
说到记忆性,那就不得不提到一个重要的问题——持续时间。体外研究发现,单核细胞/巨噬细胞的长期记忆能持续数天[6,19],其作用可持续数周[6,20]。研究显示,接种BCG的志愿者的循环单核细胞,其训练免疫能持续3个月甚至1年[21],BCG或麻疹疫苗对机体的非特异性保护可以持续数月甚至数年[22]。然而,单核细胞在循环系统中寿命很短,最长不超过1 d[23],寿命如此短暂的固有免疫细胞是怎样维持长期的训练免疫作用的呢?研究发现,固有免疫记忆可以通过造血干细胞和祖细胞进行传递,将对Toll样受体2(Toll-like receptor 2,TLR2)配体耐受的造血干细胞和祖细胞分化而来的巨噬细胞移植到辐射小鼠体内,小鼠也表现出对TLR2配体的耐受,并且被炎症刺激后炎症因子和活性氧的产生减少[24],肠道内感染了分节丝状菌的小鼠,其骨髓祖细胞会发生长期的功能改变,例如IL-23的分泌增加,由这种祖细胞分化而来的树突状细胞可以帮助宿主抵御溶组织内阿米巴的感染[25]。以上研究表明,训练免疫过程中,在骨髓祖细胞水平上可能就已经发生了表观遗传学重组,因此才使得这种记忆得以长期维持。
2.2转录和表观遗传机制 受过训练的固有免疫细胞的一个显著特点是在受到抗原或危险信号刺激时,能够发生比未训练细胞更为强烈的炎症基因的转录反应。这一过程由表观遗传学机制介导,通过刺激固有免疫细胞(或其前体),导致其染色质结构改变而发生基因转录差异,或引起细胞内某些微小RNA(microRNA,miRNA)水平的持续变化,造成更强的非特异性免疫反应[26]。
许多研究证明,训练免疫过程中固有免疫细胞会发生表观遗传重编程。用白色念珠菌或β-葡聚糖刺激单核细胞/巨噬细胞后,它们对后续刺激免疫的反应增强,这一功能的变化伴随着表观遗传重编程,如H3K4me1和H3K27ac增加[19,27]。LPS 和β-葡聚糖都能通过丝裂原活化蛋白激酶(Mitogen activated protein kinase,MAPK)依赖性通路,引起环腺苷酸依赖性转录因子7(Cyclic AMP-dependent transcription factor-7,ATF7)的磷酸化,降低抑制性组蛋白标记H3K9me2的甲基化水平,从而诱导训练免疫[28]。另有研究表明,用巨细胞病毒(Cytomegalovirus,CMV)诱导的NK细胞记忆,也在一定程度上与表观遗传重编程导致的前髓细胞性白血病锌指(Promyelocytic leukemia zinc finger,PLZF)和脾酪酸激酶(Spleen tyrosine kinase,SYK)表达降低有关[7]。此外,BCG疫苗能够提高健康人单核细胞产生炎症介质的水平,其机制也与表观遗传重编程导致的基因活化有关[29]。
那么表观遗传重编程是怎样影响固有免疫细胞产生训练表型的呢?人们发现,初次刺激引起的启动子和增强子上的组蛋白修饰可能会影响到第二次刺激时的免疫应答。初次刺激时,基因转录发生活化,伴随特殊的染色质标记;刺激消除后,这些标记部分脱落,残留的组蛋白标记,例如H3K4me1,会使得所标记的基因在接受第二次刺激时转录增强,使细胞表现出更强烈的免疫应答,即表现为训练免疫表型[19]。
miRNA也参与了训练免疫的形成[30],其半衰期很长,在第一次刺激后能持续存在。髓系细胞受到炎症刺激后miRNA-155发生上调,造成这些髓系细胞的过度活化[31]。推测髓系细胞持续表达miRNA-155可能会使自身处于一种敏感状态,在遇到第二次相同强度的刺激时,这些细胞会发生比第一次更为强烈的免疫反应。
上述阐述了表观遗传学在介导训练免疫方面的作用,但仍有一个关键问题亟待解决:细胞内到底发生了什么样的变化,能诱导表观遗传学发生改变并稳定存在?针对这一问题人们发现,细胞代谢与之关系密切,某些细胞代谢物是表观遗传酶的辅助因子,其浓度的改变能够影响基因转录的表观遗传学调控。下面我们将介绍代谢机制对训练免疫的影响。
2.3代谢机制 经过训练的固有免疫细胞内部代谢过程也发生了改变,这些变化与表观遗传的联系紧密[32],是训练免疫表型形成的重要组成部分。
免疫细胞在活化时会引起多条代谢途径的变化,例如M1型和M2型巨噬细胞的代谢途径就有诸多不同[33,34]。这提示我们,训练免疫过程中单核细胞/巨噬细胞发生的功能长期改变可能也受细胞代谢变化的影响。近期研究发现,β-葡聚糖诱导的训练免疫中,Akt/mTOR/HIF-1α通路介导的氧化磷酸化向糖酵解的转变发挥了重要作用[28,35]。三羧酸循环(Tricarboxylic acid cycle,TAC)代谢产物α-酮戊二酸和琥珀酸在控制两类表观遗传修饰酶(赖氨酸去甲基酶的JmjC家族和甲基-胞嘧啶羟化酶的TET家族)的活化过程中起着决定性作用[28,36],这些酶需要α-酮戊二酸作为辅助因子,而琥珀酸则会限制他们的活性。巨噬细胞产生训练免疫的过程中,胞内琥珀酸水平可能会升高,而琥珀酸可以抑制组蛋白去甲基化酶JMJD3的活性,导致特定基因(例如与M2表型相关基因)的H3K27三甲基化增强,从而抑制它们的表达[37],使得训练巨噬细胞表达促炎表型,并保持到下一次刺激来临。另外,乙酰CoA的积累会引起组蛋白乙酰化[27],延胡索酸盐(TAC中间产物之一)的积累可以抑制组蛋白去甲基化酶的活性[38],诱导胆固醇合成导致甲羟戊酸积累,也能引起训练免疫中的表观遗传重编程[39]。当糖酵解被抑制,表观遗传重编程所造成的训练免疫也会被抑制[40,41]。上述证据表明,代谢过程的确参与了训练免疫的形成。目前有两个问题仍需解决:解释这些代谢物潜在的非特异性功能是如何具有位点或基因特异性效应的,以及寻找训练免疫过程中代谢物影响DNA和组蛋白修饰酶活性的直接证据。
3 训练免疫在疾病方面的研究进展
如前所述,训练免疫能使机体对刺激产生更为强烈而持久的免疫反应,来抵抗再次感染。诱导训练免疫或许可以治疗某些固有免疫反应功能缺陷性疾病,但异常活化的训练免疫也可能会导致过度的炎症反应,对机体造成损害。在一些慢性炎症状态下,由内源性抗原诱导的训练免疫会加剧组织损伤。下面我们将对训练免疫在自身免疫性疾病和自身炎症性疾病中的潜在作用进行讨论。
3.1类风湿性关节炎 类风湿性关节炎(Rheumatoid arthritis,RA)是最常见的炎症性关节炎之一,其病理过程非常复杂,疾病最终会导致滑膜炎症和骨与软骨的破坏[42]。固有免疫系统在RA的发生发展中发挥重要作用,固有免疫细胞也能造成组织损伤性炎性病变[43]。RA患者的循环单核细胞中,多种促炎因子基因的表达出现上调[44,45],这与具有训练免疫表型的单核细胞相类似;另外,其PI3K/mTOR和MAPK这两种参与训练免疫的信号通路也发生了活化[45,46],抑制RA患者体内mTOR信号通路的表达可以减少其滑膜破骨细胞形成并防止局部骨侵蚀和软骨丢失,从而缓解疾病[47,48]。
有研究发现表观遗传重编程在RA中发挥一定的作用。RA患者滑膜组织中的组蛋白乙酰基转移酶(Histone acetyltransferase,HAT)/组蛋白去乙酰化酶(Histone deacetylase,HDAC)平衡向组蛋白乙酰化的方向移动[49];阿达木单抗和依那西普会下调巨噬细胞中H3K4、H3K27、H3K36和H3K79的三甲基化水平以及细胞趋化因子CCL-2 (MCP-1)启动子位点的组蛋白3和4的乙酰化水平,这些表观遗传学修饰都与RA发病有关[50]。
训练免疫中发生的代谢改变同样也出现在RA患者巨噬细胞内。LPS刺激下,RA患者巨噬细胞内腺苷三磷酸(Adenosine triphosphate,ATP)水平更高[51],糖酵解中的限速酶,丙酮酸激酶同工酶 2(Pyruvate kinase isozymes M2,PKM2)、6-磷酸果糖2激酶/果糖-2,6-二磷酸酶 3(6-Phosphofructo-2-kinase/Fructose-2,6-biphosphatase 3,PFKFB3)和己糖激酶 2(Hexokinase 2,HK2)以及葡萄糖转运体1(Glucose transporter 1,GLUT1)和GLUT3活性增强,葡萄糖摄取和氧的消耗增加[52],谷氨酸盐、琥珀酸盐和延胡索酸盐累积[40,42]。这些都标志着RA促炎巨噬细胞中出现了与训练免疫类似的代谢状态[53,54]。
由此可见,RA患者的固有免疫细胞具有许多训练免疫的特点。研究发现,关节局部无菌性炎症及组织损伤可以释放DAMPs,并诱导训练免疫[55],但RA中训练免疫究竟是如何被诱导产生的还未可知,具体机制亟待后续的研究证实。
3.2系统性红斑狼疮 系统性红斑狼疮(Systemic lupus erythematosus,SLE)的发病伴随多种免疫系统异常,最广为人知的病因是抗核抗体的大量产生。目前发现SLE患者的单核细胞发生了表观遗传重编程,其TNF-α基因区周围的组蛋白高度乙酰化,使得该段基因更容易进行转录[56],组蛋白H4为高度乙酰化[57],同时也检测到了SLE特异性改变H3K4me3和H3K27me3的增强子[58]。对SLE患者的原代单核细胞进行全基因组表观遗传分析发现,炎症和免疫应答相关基因表现为高度H3K4me3,这与β-葡聚糖诱导的单核细胞中H3K4me3的情况相类似[59,60]。
SLE患者体内,免疫细胞的代谢也发生了变化,并可能对表观遗传学造成影响[61]。训练免疫中mTOR通路发生活化。在SLE的许多免疫细胞包括单核细胞当中,mTOR通路也都处于活化状态[62-64],雷帕霉素抑制mTOR能有效缓解疾病[62]。这一系列证据都表明了SLE患者体内可能存在训练免疫。
3.3自身炎症性疾病 自身炎症性疾病是一组由固有免疫系统介导的免疫系统疾病,特点表现为周期性发作的发热和炎症,常见症状有关节疼痛、皮疹、腹痛,长期可导致淀粉样病变。
这些自身炎症性疾病的共同特点是IL-1β的产生过剩,因此抗IL-1疗法能够用于治疗这类疾病[65]。IL-1β是维持疾病的重要因素,同时也是训练免疫重要的诱导剂。研究表示,给小鼠注射IL-1β可以降低随后细菌和真菌感染所导致的死亡率[66,67]。这意味着IL-1β能诱导单核细胞和巨噬细胞发生表观遗传重编程,从而产生更多的促炎免疫细胞,构成训练免疫,保护机体免遭后续感染。对人类单核细胞进行体外训练结果发现,用IL-1β训练的单核细胞在LPS刺激下能够产生更多的IL-6 和 TNF-α,且IL-6和TNF-α启动子的H3K4me3也增多[10]。IL-1能够通过诱导训练免疫抵御感染,但也可以导致过度炎症,后者可能会引发自身炎症性疾病,关于这一点还需要更深入的研究。
3.4干燥综合征 干燥综合征(Sjogren syndrome,SS)是一种慢性自身免疫性疾病,表现为唾液和泪腺功能障碍。研究表明,SS中的巨噬细胞具有训练免疫表型。SS患者外周CD14+单核细胞受到凋亡细胞刺激时,其TNF-α或IL-1β的产生量会增加,IL-10产生量减少,表现为促炎[68,69];单核细胞来源的树突状细胞受到LPS刺激时也表现为促炎,TNF-α、IFN-γ诱导单核因子(Monokine induced by IFN-γ,MIG)、干扰素-α(Interferon-α,IFN-α)、IL-6、IL-12、单核细胞炎性肽(Monocytic inflammatory peptide,MIP)-1α/β、MCP-1的表达水平都高于健康人[70],SS患者循环系统中的未受到刺激的单核细胞内部,Ⅰ型IFN相关基因的表达高于健康人的单核细胞[71-73],单核细胞被IFN-γ刺激后,IL-6和B细胞活化因子(B cell activating factor,BAFF)的产生也增多[74]。
目前发现SS患者的外周血单个核细胞(Peripheral blood mononuclear cells,PBMCs)中miRNA-146a和181a表达上调,单核细胞中miRNA-34b-3p、miRNA-4701-5p、miRNA-609、miRNA-300、miRNA-3162-3p和miRNA-877-3p表达上调[75,76]。这些miRNAs都能够抑制转化生长因子-β(Transforming growth factor-β,TGF-β)信号通路,激活IL-12和TLR/NF-κB信号通路[77]。mTOR抑制剂雷帕霉素纳米粒可以用于SS小鼠模型的治疗,减少泪腺损伤[78],而mTOR通路的活化也是产生训练免疫的关键机制之一。
3.5白塞病综合征 白塞病综合征(Behcet′s disease,BD)是一种炎症性疾病,表现为口腔和生殖器病变、关节炎和葡萄膜炎,病因未知。BD患者的外周单核细胞处于活化状态,产生促炎因子增加;CD14+单核细胞在IFN-γ刺激下产生趋化因子CXC配体-9(Chemokine CXC ligand-9,CXCL-9)和CXCL-10的量高于健康人[79],说明BD患者的单核细胞可能存在训练免疫表型。BD患者单核细胞的全基因组DNA甲基化情况显示,383 个CpG 位点(胞嘧啶-磷酸-鸟嘌呤位点,即DNA序列中胞嘧啶后紧连鸟嘌呤的位点)与健康人不同(CD4+T细胞中只有123个不同),可能存在于训练免疫有关的表观遗传重编程现象[80]。
综上所述,目前发现训练免疫能够参与自身免疫性疾病和自身炎症性疾病的发生、维持和恶化。在疾病发病阶段,基因或环境因素(或二者的结合)会“初步训练”单核细胞/巨噬细胞,从而引发疾病。在疾病发展阶段,经过训练的单核细胞/巨噬细胞更容易活化,从而导致症状的维持或恶化。因此,针对疾病不同阶段的研究,应当考虑上述差异而采用不同的造模方式,从而更加准确地研究训练免疫在其中的潜在作用。通过药物等手段来调整训练免疫中的表观遗传重编程或许可以使固有免疫系统恢复正常平衡,从而起到疾病治疗的作用。
4 结论及展望
训练免疫是一个崭新的概念,是哺乳动物免疫反应中宿主防御的一种基本属性,它的提出为人类免疫反应的研究提供了重要的信息。这个令人兴奋的新领域还有许多研究空间:①揭示介导训练免疫的分子机制以及免疫、代谢、表观遗传学过程,阐明训练免疫的持续时间以及它对髓系祖细胞和组织巨噬细胞的影响;②运用尖端技术,如单细胞转录组和表观基因组学(尤其是DNA甲基化),寻找新的具有固有免疫记忆特征的细胞亚群,开辟针对特定细胞亚群的新疗法;③挖掘训练免疫在宿主防御受损性疾病(如脓毒症后免疫麻痹或癌症)以及自身炎症性疾病和自身免疫病当中的作用;④固有免疫记忆的概念可以用于协助设计新的治疗方法,尤其是那些能够摧毁宿主防御的疾病,如脓毒症中的免疫麻痹或肿瘤,或者那些训练免疫被异常激活的疾病,如自身免疫性疾病。
