反刍动物POU1F1基因多态性与经济性状关联研究进展
2019-05-27张阳海潘传英雷初朝蓝贤勇
杨 韩, 张阳海, 潘传英, 雷初朝, 陈 宏, 蓝贤勇
(西北农林科技大学动物科技学院,陕西 杨凌 712100)
垂体特异性转录因子(pituitary specific transcription factor,POU1F1,又叫PIT-1或GHF-1)是POU(Pit-Oct-Unc)家族成员之一,在垂体前叶特异性表达,是重要的组织特异性转录因子[1]。POU1F1具有高度保守的DNA结合结构域,通过识别并结合催乳素(prolactin,PRL)、生长激素(growth hormone,GH)和促甲状腺素β亚基(thyroid-stimulating hormone beta subunit,TSHβ)基因启动子中特异的DNA序列,对这些基因起正向调控作用,从而对哺乳动物生长发育等方面起着重要作用[2]。研究表明,POU1F1基因突变会导致垂体发育不全,造成动物体内PRL、GH、TSH激素的缺乏,从而阻碍动物生长发育。在小鼠上,POU1F1基因突变导致Snell矮小型(dw)和Jackson矮小型(dw-J)出现[3];同时,人POU1F1基因突变会导致联合性垂体激素缺乏症(combined pituitary hormone deficiency,CPHD),患者发育迟缓,生长出现明显障碍[4]。
此外,有大量研究报道了POU1F1基因多态性与牛、山羊、绵羊等反刍动物的生长性状之间存在显著关联[5-7]。以上研究表明POU1F1基因对于反刍动物生长、繁殖及泌乳等具有重要的调节作用,据此,本文主要就牛羊等反刍动物POU1F1基因结构及生物学特性、基因多态性及其与经济性状关联研究等进行综述。
1 反刍动物POU1F1基因的结构特征及其生物学特性
牛、绵羊和山羊POU1F1基因均定位于1号染色体的q21-22区[8]。该基因结构高度保守,一般由6个外显子,5个内含子及其他相关DNA元件组成(图1)。POU1F1基因表达于动物的垂体前叶细胞,其表达产物POU1F1蛋白包含3个重要的功能区,其一是含129个氨基酸的N端区,其二是含142个氨基酸的POU蛋白区,其三为含20个氨基酸的C端区;其中POU结构域的主要功能是识别DNA特异顺序,并与之呈高亲和力结合,从而启动靶基因转录,该区又可分为POU同源区(POU-homeo domain,POU-HD)及POU特异区(POU-specific domain,POU-SD)两部分[9]。
POU1F1是垂体生长激素(GH)细胞,催乳素(PRL)细胞以及促甲状腺素(TSH)细胞内基因转录的重要调节者,能特异地识别基因上的DNA序列,并与之结合,进而影响相关基因转录,最终影响家畜生长、繁殖及泌乳等各个方面。据此,该基因的多态性及其在家畜育种工作上应用研究成为国内外诸多学者研究重点。国内外研究表明,POU1F1基因多态性对牛的体重、产奶量等生产性状有显著影响[10-11],对羊的产羔数、体重、产奶量、产绒量等性状有显著影响[12-15]。因此,对POU1F1基因调控机制的深入研究对于提高牛羊等反刍动物生产成绩至关重要。
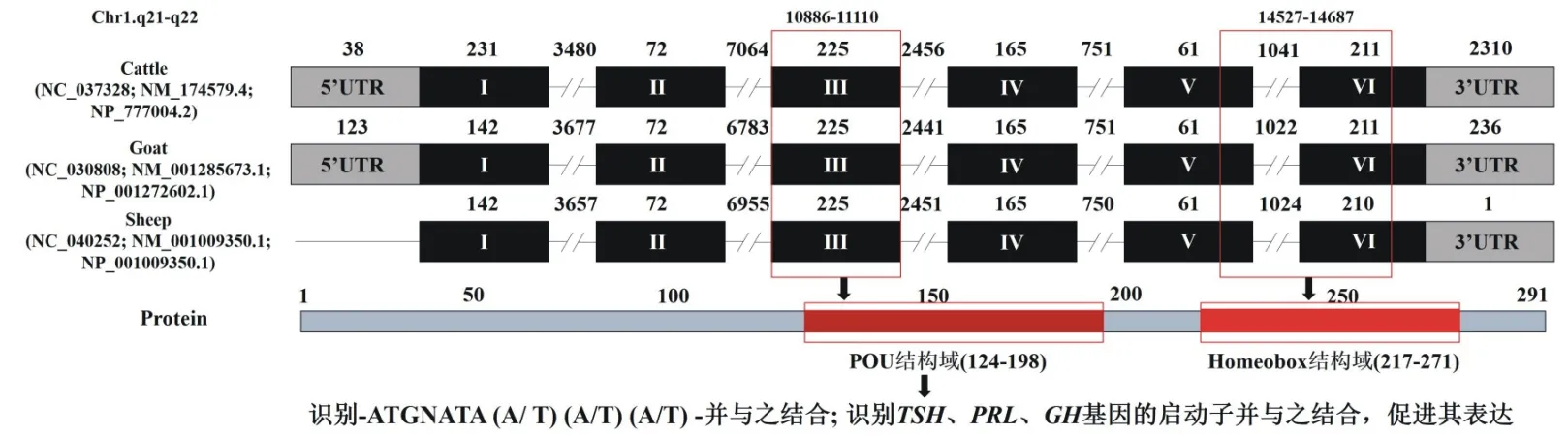
图1 反刍动物POU1F1基因结构特征及其生物学特性示意图
2 反刍动物POU1F1基因多态性研究进展
2.1 牛POU1F1基因多态性与经济性状的关联研究
此前国内外许多学者以POU1F1基因作为牛生产性状选育改良的候选基因之一,围绕基因多态位点与经济性状的关系开展了大量研究。20世纪90年代,国外最先开始对牛POU1F1基因多态性进行挖掘。1994年,Woollard等[16]首次报道牛POU1F1基因第6外显子遗传变异位点,发现了可被HinfI内切酶识别的SNP位点。2000年,Woollard等[8]进一步将牛POU1F1基因定位于lq21-22区域,为进一步探究该基因对牛生产性状的影响打下基础。最近于2018年,Pytlewski等[17]在Holstein-Friesian牛POU1F1基因第6外显子上扫描到1个SNP位点(c.1178G>A),统计分析结果表明该位点与体重、初配年龄、产后休息期、受胎率、产犊后奶牛体重、初生重等显著相关(P<0.05)。同年,Thuy等[18]在Holstein-Friesian牛第2外显子和第3外显子上分别检测到SNP位点c.545G>A和c.577A>C;相关性分析表明,c.545G>A位点与产奶量显著相关(P<0.05),而c.577A>C位点则不存在相关性(P>0.05)。目前为止,关于POU1F1基因的研究在牛上做了包括Fleckvieh牛、Montbeliarde牛、南阳牛、秦川牛、郏县红牛、中国荷斯坦牛、鲁西牛、安格斯牛、晋南牛、固原牛、Holstein-Friesian牛、Nellore牛、Hanwoo牛、Canchim牛、Zebuine牛、皮埃蒙特牛、新疆褐牛、申扎、类乌奇、斯布、帕里共21个品种,共检测了10个SNP位点与牛的体重、体高、体长、体斜长、胸围、腹围、产奶量、乳蛋白含量、乳脂含量、生产寿命、背膘厚、初生重、断奶前平均日增重、料肉比、胸深、胸宽、初配年龄、产后休息期、受胎率、产犊后母牛体重等20个性状相关联,其中大部分研究集中于POU1F1基因第6外显子区域多态性(表1)。根据现有研究,发现位于第6外显子上的HinfI位点在各个品种牛的基因组上均广泛存在,且与多个生长性状及泌乳性状相关联,提示该位点可作为潜在的分子标记进行优良牛种选育。另外,目前的研究多集中于奶牛、黄牛等常见牛种上,而属于青藏高原及其毗邻高寒牧区特有的牦牛上研究却较少。作为高寒地区重要的品种,应加强对POU1F1基因多态性的研究,对提高牦牛的生产性能具有重要意义。
2.2 山羊POU1F1基因遗传变异与经济性状的关联研究
2000年,Woollard等[8]将山羊POU1F1基因定位于1q21-22区域。在此基础上,国内最先就山羊POU1F1基因多态性及其与生产性状的关系开展了大量研究,而国外对此鲜有报道。2007年,Lan等[12-13,32-33]首次报道山羊POU1F1基因外显子6及其3′-UTR区域多态性(AluI、PstI和DdeI位点)对产奶量、产羔数和产绒性状等生产性状的影响。2009年,Lan等[34]对内蒙古白绒山羊群体中POU1F1基因上未报道的外显子1~5和5′UTR区域的遗传变异情况进行了筛选,共发现12个SNP位点(表2);进一步分析表明,这些多态位点对产绒量和羊绒纤维长度有显著的影响。2012年,Feng等[7]在5个中国本地山羊品种上检测到6个突变,包括第3外显子C256T、第3内含子C53T和T123G,第6外显子G682T(A228S)、T723G和C837T,分析结果初步表明济宁灰山羊POU1F1基因C256T和G682T位点的T等位基因与高产羔数之间存在显著关联。最近,Zhu等[15]在陕北白绒山羊中检测了4个SNP位点,其中c.682G>T、c.723T>G和c.837T>C位于POU1F1基因第6外显子,均表现为3种基因型;而定位于3′-UTR区域的c.876+110T>C在该群体中未分型;进一步分析发现,c.682G>T和c.837T>C位点与产羔数显著相关(P<0.05),c.682G>T、c.723T>G和c.837T>C位点对生长性状有显著影响(P<0.05);双倍型分析表明,不同的双倍型和产羔数及生长性状之间存在强相关性,且具有H3H7双倍型(GTTT-TTTT)的雌性山羊比其他双倍型个体拥有更多的产羔数和更优的生长状况(P<0.05)。截至目前,关于山羊POU1F1基因多态性研究主要集中于内蒙古白绒山羊、济宁灰山羊、贵州白山羊等15个中国本地山羊品种,共检测到25个SNP位点,与绒、肉、奶等13种生产性状显著相关(表2)。

表1 牛POU1F1基因单核苷酸多态性汇总
注:E为外显子(exon);I为内含子(intron)。下同。
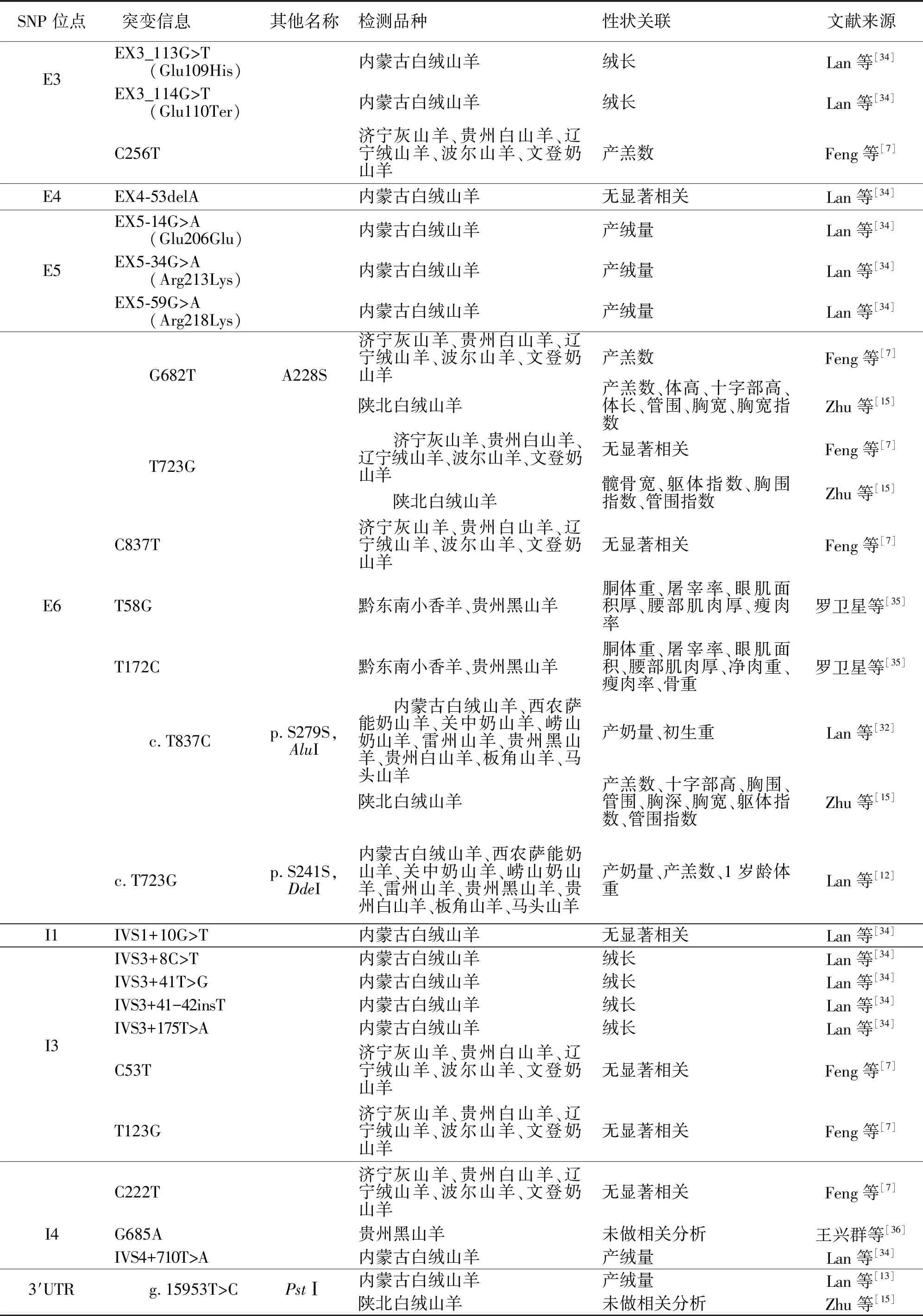
表2 山羊POU1F1基因单核苷酸多态性汇总
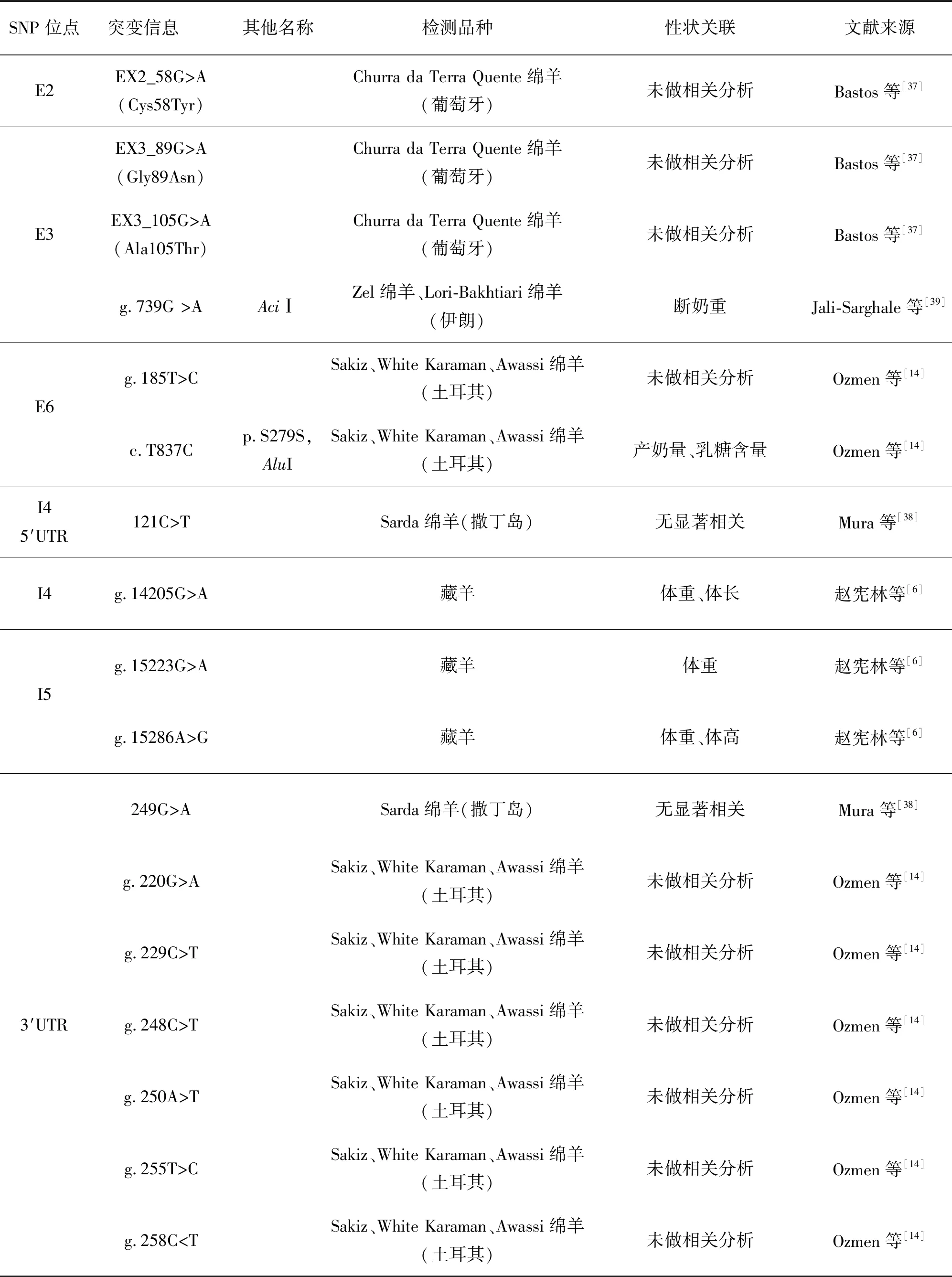
表3 绵羊POU1F1基因单核苷酸多态性汇总
2.3 绵羊POU1F1基因遗传变异与经济性状的关联研究
在绵羊上,国内外关于POU1F1基因遗传变异的研究工作较少。2006年,Bastos等[37]首次报道了绵羊POU1F1基因的结构,包含6个外显子和5个内含子,长度为5 787 bp;同时发现了葡萄牙本地绵羊POU1F1基因第2、3外显子上存在三处错义突变,这些突变可能改变POU1F1蛋白的结构和活性;同时,在第4内含子上也发现存在一处突变,但未在外显子1、4和6中检测到突变。2017年,赵宪林等[6]在藏羊POU1F1基因第4内含子上检测到1个SNP位点(g.14205G>A),在第5内含子上检测出2个SNP位点(g.15223G>A和g.15286A>G);关联性分析表明,g.14205G>A位点与藏羊体质量和体长显著相关(P<0.05),AA基因型个体为优势个体;g.15223G>A位点与藏羊体质量显著相关(P<0.05),GG基因型为优势基因型;g.15286A>G位点与藏羊体质量和体高显著相关(P<0.05),AA基因型个体为优势个体。从2006年第1篇报道到现在为止,关于绵羊POU1F1基因的研究仅对包括Churra da Terra Quente绵羊(葡萄牙)、Zel绵羊(伊朗)、Lori-Bakhtiari绵羊(伊朗)、Sakiz(土耳其)、White Karaman(土耳其)、Awassi绵羊(土耳其)、Sarda绵羊(撒丁岛)、藏羊在内的8个绵羊品种进行了检测,发现了17个SNP位点,其与断奶重、产奶量、乳糖含量、体重、体长、体高等生产性状显著相关(表3)。这些结果表明绵羊POU1F1基因上存在较为丰富的多态性,应当大力发掘,以期寻找与绵羊优良生产性状相关的分子标记,为绵羊的选育改良提供基础。
3 其他家畜POU1F1基因遗传变异与经济性状的关联研究
鉴于POU1F1基因对动物生长发育起着十分重要的作用,除了对牛、山羊、绵羊等反刍动物POU1F1基因多态性进行研究以外,在猪、马等家畜及家禽上也开展了大量研究。2011年,宋成义等[40]在猪POU1F1基因的1.5 kb启动子区域发现了5个突变位点,其中5 bp插入/缺失突变A等位基因(128 bp/42 bp)的频率在引入品种猪中最高,在培育品种苏姜猪中处于中等,而在中国地方品种最低;进一步研究发现该突变与猪6月龄的体高、体长、胸围和体重等生长性状显著相关(P<0.05)。2013年,Yan等[41]在POU1F1基因中检测到1个SNP位点(NC_006088.2:g.3109G>A),相关性分析表明该位点的GA基因型个体在16周时体重较大,300日龄时所产生的卵子数较多(P<0.05)。2017年,于农淇等[42]发现伊犁马POU1F1基因内含子区的SNP(g.2758G>A)与日均母马产奶量差异不显著(P>0.05)。2018年,徐珑洋等[43]在藏鸡、沪宁鸡POU1F1基因的外显子2上发现1个多态位点(C/T),其中C等位基因为优势等位基因,而CT基因型是控制泸宁鸡早期生长速度的优势基因型,因此该位点可以作为影响鸡早期生长速度的1个潜在分子标记进行选育。
4 展 望
POU1F1基因被认为是神经—内分泌—生长轴的重要成员之一,其通过调控TSH、PRL、GH基因表达及垂体细胞的分化,对动物生长发育及繁殖等起重要调节作用。目前,国内外对牛和山羊,以及猪、家禽等家养动物POU1F1基因及其对生长发育的影响的相关研究越来越多,但对绵羊和牦牛的关注度有所不足,遗传效应研究较少。综上所述,对反刍动物POU1F1基因的深入研究,对畜禽的选育改良具有很重要的理论价值和实际意义。
