模拟增温对杉木幼树生长和光合特性的影响
2019-05-13叶旺敏熊德成杨智杰朱益广张秋芳刘小飞林伟盛杨玉盛
叶旺敏,熊德成,*,杨智杰,朱益广,张秋芳,刘小飞,林伟盛,胥 超,张 景,杨玉盛
1 福建师范大学地理科学学院, 福州 350007 2 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 3 宾夕法尼亚州立大学, 帕克 PA 16802
森林作为全球陆地生态系统的主体,是地球生物圈的重要组成部分,较其他植被类型相比,森林植被具有分布广、生产力高及生物量大等特点,在陆地碳循环中发挥着重要作用[1- 2]。据联合国气候变化专门委员会第五次气候变化评估报告,预计到本世纪末全球地表平均增温0.3—4.8℃[3]。气候变暖导致气温和土壤温度升高,必将对森林植被造成剧烈影响。光合作用作为植物行为最重要的生理过程,极易受到温度变化的影响[4- 5]。温度升高可以引起植物叶片形态结构、气孔导度、光合色素含量、以及植物进行光合作用所需的一系列酶的活性等发生改变,从而影响光合电子的传递速率,改变植物的光合效率[6- 8]。国内外关于植被光合能力对增温响应机理的报道已有很多,但由于增温方式、区域及种类的不同,并没有得出一致的结论。如任洁等[9]采用气候生长箱对银中杨设置3个温度梯度处理发现,杨树树皮绿色组织和叶片的光合能力随着温度的升高显著增强;韩超等[10]采用开顶式增温方法对云杉进行测定,结果表明增温促进云杉叶片叶绿素含量合成,进而提高其光合能力;另有研究表明增温抑制植物光合作用[11- 13];少数研究则认为增温对植物光合速率无显著影响[14]。另外,大部分研究结果还表明,当植物所处的环境温度低于植物生长最适温度时,温度升高对植物生长和光合效率有益,当温度超过植物生长最适温度时对植物生长、光合效率和生产力起负效应[15- 17]。
纵观过去20年,大量模拟增温控制实验相继开展,但大多数实验主要集中在高纬度地区农田、冻原、草原及森林生态系统,有关低纬度地区模拟增温实验仍鲜见报道[18- 20]。由于研究的缺乏,低纬度地区森林植被对未来全球气候变暖的响应仍存在许多不确定性。尽管长期以来的观点认为低纬度地区变温幅度可能远小于高纬度地区[21],但越来越多的研究已经证实,相比于高纬度地区,低纬度地区森林植被对热适应的生态位相对较窄,其适应生态位温度范围以外的温度的能力较弱,对未来全球气候变化的适应能力可能远小于高纬度地区[22- 23]。低纬度森林植被具有丰富的碳储量,森林生产力较高,在我国陆地生态系统碳循环中占有重要地位[24],光合作用作为植物物质生产的基础,是决定森林生产力的主要因素,也是森林生态系统碳收入的唯一途径和碳循环开始的重要环节。温度通过改变植物光合能力的大小,进一步影响植物的生长生理过程,在未来全球气候变暖大背景下可能会对亚热带地区森林植被造成剧烈影响。因此,研究低纬度地区森林植被光合能力与气候变暖之间的相互关系及其响应与适应对策,对低纬度森林植被演化和预测其在未来生境下变化特征具有重要意义。
杉木(Cunninghamialanceolata)作为亚热带地区主要造林树种之一,广泛分布于我国南方16个省区,面积占世界人工林面积的6%,我国面积的19%、蓄积量的25%,由于面积较广,碳汇能力较强,杉木人工林可以部分反映出我国人工林碳汇变化情况,在我国人工林生产中占有重要地位[25- 26]。为此,本研究选取杉木为研究对象,通过野外模拟土壤增温实验,测定土壤增温对杉木幼树生长、叶片光合作用参数的影响,并通过测定土壤,叶片养分特征,叶片叶绿素含量以及非结构性碳水化合物含量,进一步解释杉木生长特征及光合能力对增温的响应机理。实验的开展将揭示亚热带杉木生长特征和光合能力对全球变暖的适应性,并为预测未来我国亚热带人工林植被碳汇供给潜力提供参考依据。
1 材料与方法
1.1 样地基本概况
本试验地点位于福建省三明市森林生态系统与全球变化研究站—陈大观测点(26°19′N,117°36′E),该地区平均海拔300 m,年均气温19.1℃,年均降水量1749 mm,年均蒸发量1580 mm,相对湿度81%,属中亚热带季风气候。土壤为黑云母花岗岩发育的红壤。
1.2 实验设计
增温平台设置实验小区面积为2.5 m×2.5 m,土壤深度为50 cm,样地中土壤均取自观测站附近杉木人工林的土壤(0—40 cm),填土过程主要是将取回的土壤去除其中的根系和沙石,充分混合均匀后分别填入建好的实验地中,以此尽可能的消除土壤和样地间的异质性。实验设置增温(CT)和对照(W)2种处理,每种处理共6个重复,共计12个小区。于2014年10月份在各个实验小区地表下10 cm处平行布设加热电缆以及一个土壤温、湿度传感器,电缆功率为40 W/m2,间距为20 cm,并在最外围环绕一圈,以确保样地增温的均匀性,传感器用来监测土壤温、湿度的变化情况。2015年1月在各个小区种植5棵生长情况大致相同的一年生三代杉木幼苗(生长特征参数见表1),每株杉木种植在两条电缆线中间位置。全年不间断的持续进行土壤增温,增温幅度主要是指增温处理各个小区10 cm的平均土壤温度较对照增加4℃。

表1 不同处理杉木初始生长特征参数
1.3 杉木生长及叶片光合作用参数测定
于2016 年3—5月每半个月对两种处理的所有杉木进行生长量测定,测定指标主要包括杉木的树高、地径。
2016年5月底,选择晴朗天气,在8:30—11:00,采用CIRAS- 3便携式光合测定仪(英国)原位测定杉木叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)和水分利用效率(WUE)。设定人工光源(红蓝光源),叶室环境因子条件设置为:叶片温度27℃,相对湿度50%—65%,光强度为1500 μmol m-2s-1,大气中CO2浓度为380 mol/mol。测量期间,每个重复选取三片中部当年生健康向阳叶片进行测定。由于杉木幼苗叶片细长,CIRAS- 3叶室无法完全覆盖叶片面积,故采取标记方法,每测1片,标记所测叶片面积宽度,每个重复共取3片,标记完,用剪刀剪去未测量到的叶片部分。利用扫描仪测定实际光合测量面积,通过计算得出该部分叶片的实际光合参数。
1.4 叶绿素含量和非结构性碳水化合物的测定
光合参数测定完毕后,对每个处理所有重复相同部位叶片均匀采样,叶片本身为健康无病害的当年生鲜叶,分别装入自封袋,标记,将每袋自封袋叶片分装成两袋,一袋叶片直接保存在放有冰袋的保鲜箱中,同一天带回实验室进行叶绿素含量测定。另一部分进行烘干,并利用球磨仪进行粉碎,装入自封袋,供NSC含量测定。叶绿素含量测定参照王学奎等[27]采用80%丙酮研磨法,具体操作如下:先将每包叶片剪碎混合均匀,称取混合后叶片0.5 g,置于研钵中,添加些许石英砂和碳酸钙粉末以及5 mL纯丙酮后开始研磨至匀浆,再加入5 mL 80%的丙酮溶液,继续研磨至组织变白,后转入离心管进行离心,离心完毕,取离心管上清液于20 mL试管中,用80%丙酮定容。再用津岛-UV2450紫外分光光度计测量663 nm、645 nm吸光度,根据公式计算叶绿素含量浓度。
叶片非结构性碳水化合物(NSC)含量方法参照Bussy等[28]、于敏丽等[29]采用酚-浓硫酸法(NSC含量为测得淀粉含量和可溶性糖含量之和)。
1.5 土壤有效氮、叶片氮含量测定
于2016年5月底同步在两种处理的各个重复用直径2 cm土钻对0—10 cm土壤进行取样,装袋,用冰袋进行保存并立即带回实验室进行处理。去除土壤中根系和砂砾,过2 mm筛,装入自封袋备用。土壤铵态氮和硝态氮用2 mol/L的KCl溶液按5∶1液土比浸提,然后用连续流动分析仪(Skalar san++,Skalar,荷兰)进行测定。叶片总氮含量采用碳氮元素分析仪(Elementar Vario MAX,Elementar,德国)进行测定。
1.6 数据处理
利用Excel 2003对所有数据进行平均值和标准误处理计算,使用统计软件SPSS 20.0中单因素方差分析比较分析,对增温和对照处理杉木生长、光合作用参数、土壤有效氮,叶片总碳、总氮含量,叶绿素含量和非结构性碳水化合物指标进行显著性分析,显著性水平设定为P=0.05。相关图表采用Origin 9.0完成。
2 结果和分析
2.1 增温对土壤温度和含水量的影响
2016年3—5月份,增温处理土壤温度平均值为24.6℃,对照平均温度为20.42℃,增温后,与对照相比,增温样地的土壤温度平均升高了(4±0.2)℃(图1)。增温样地土壤湿度平均值为19.9%,对照样地为2.67%,与对照相比,增温后土壤湿度下降了12.2%(图1)。

图1 增温和对照土壤温度和湿度动态变化Fig.1 The dynamic change of soil temperature and humidity
2.2 增温对杉木生长量和光合特性参数的影响
增温后杉木生长较对照均表现出增加趋势,但并不显著(图2)。增温和对照处理中杉木幼树树高分别为234.4,217 cm;地径分别为45.3,44.3 mm(图2)。
增温处理杉木幼树叶片的净光合速率和水分利用效率较对照显著增加(P<0.05),分别升高了71.38%和51.27%(图3)。气孔导度、蒸腾速率和胞间CO2浓度均表现出增加趋势(P>0.05),分别增加了17.1%、21.4和10.5%(图3)。
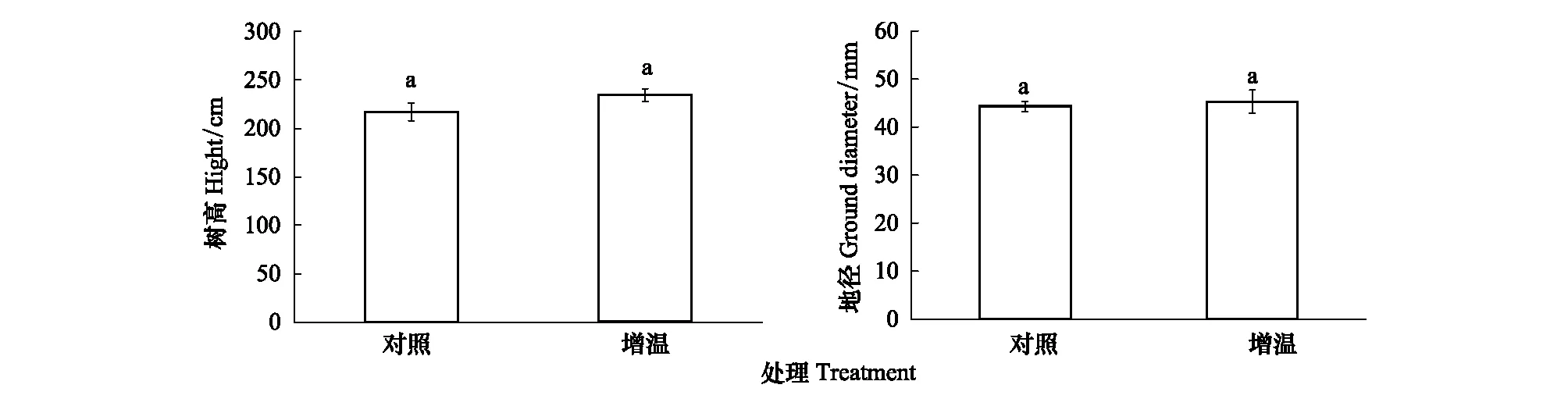
图2 增温和对照处理杉木生长特征Fig.2 The growth characteristic ofCunninghamia lanceolate
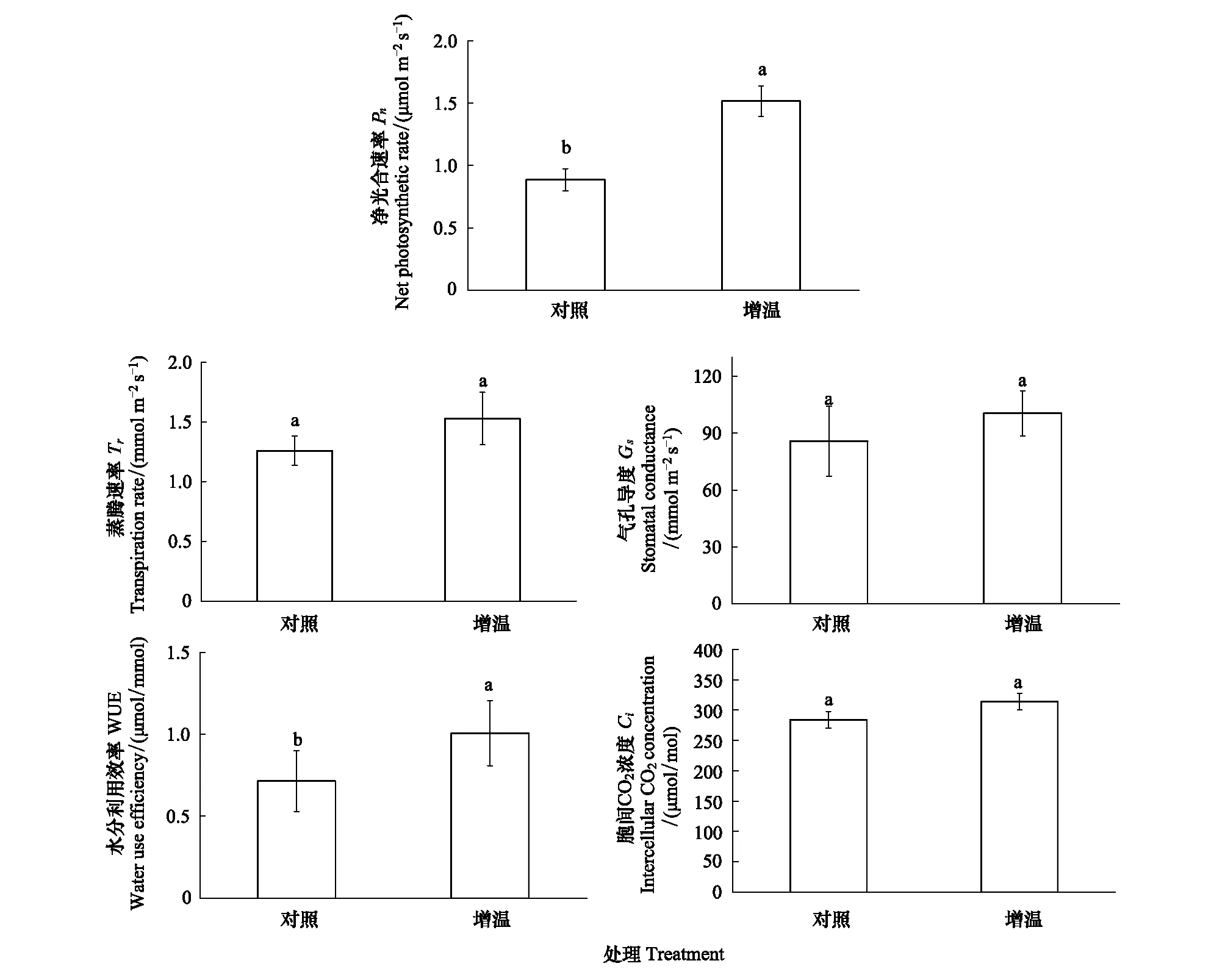
图3 增温处理下杉木叶片光合特征参数Fig.3 The photosynthetic characteristics ofCunninghamia lanceolatain control and warming treatment
2.3 增温对土壤、叶片养分的影响
森林土壤中有效氮主要以铵态氮和硝态氮的形式存在,两者可供植物直接吸收利用,是决定植物生长生理的重要养分因子。增温后,土壤中的硝态氮和铵态氮含量较对照相比显著增加(P<0.05),分别升高48.6%和16.1%(图4)。叶片氮含量作为叶绿素的重要组成物质,与叶片光合能力紧密联系。增温处理下,叶片氮含量较对照显著增加(P<0.05),增加比例为31.4%(图4)。

图4 增温处理下土壤有效氮、叶片氮含量Fig.4 The characteristics of soil and leaf nutrients in control and warming treatment
2.4 增温对叶片叶绿素含量、非结构性碳水化合物的影响
杉木幼树叶片在增温后叶绿素a、叶绿素b及叶绿素总量均显著高于对照(P<0.05)(图5),分别增加了76.3%、55.8%、68.7%,叶绿素a/b也表现出增加趋势(图5)。叶片NSC含量可以用来反映树木碳收支平衡,经计算得出,杉木幼树在增温处理下,非结构性碳水化合物含量较对照表现出增加趋势(分别为185.19 mg/g,171.57 mg/g),但与对照相比并无显著差异(P>0.05)(图5)。

图5 增温处理下叶片叶绿素含量、非结构性碳水化合物含量Fig.5 The chlorophyll content and non-structural carbohydrate content of leaves in control and warming treatment
3 讨论
3.1 土壤增温对杉木幼树叶片光合能力的影响
3.2 土壤增温对杉木幼树生长的影响
温度作为最重要的生态因子之一,对植物的生长和发育起着至关重要的作用。本研究中,尽管增温显著提高土壤养分有效性和杉木本身的光合能力,但增温对杉木幼树的生长指标(树高、基径)影响并不显著,这与以往的大多数研究结果并不一致,如石福孙等[43]认为高寒地区植被所处的环境温度普遍低于其最适温度值,因此增温可以显著提高高寒地区植被的生长速率。NSC含量的大小反映了净光合作用碳吸收与不可逆转的结构上的投资与呼吸作用消耗(汇)之间的平衡[44]。本研究发现,增温后杉木叶片NSC含量并无显著变化。因此,我们可以推测引起杉木生长无显著差异的原因可能有以下两个方面,一是目前亚热带地区本身温度较高,杉木所处的环境温度可能已接近其生长的最优温度,增温并不能显著提高杉木生长速率。二是从杉木叶片NSC含量变化趋势来看,增温亦无显著影响,说明土壤增温显著提高杉木幼树光合作用能力的同时,夜间增温引起的根际高温亦显著增加了杉木幼树的呼吸消耗,使杉木叶片NSC含量无显著差异,进而导致杉木生长无显著变化。另外,也可能在于本研究中增温时间尚短,造成杉木生长特征对增温的响应并不明显,还有待进一步观测。
4 结论
土壤增温通过气孔限制因素和非气孔限制因素显著增加杉木幼树叶片的光合能力。气孔限制因素:土壤增温使杉木幼树叶片维持较高的气孔导度,导致叶片吸收CO2能力增强,可供其进行光合作用的原料增多。非气孔限制因素:土壤增温后土壤有效氮含量显著增加,使植物体内养分状况发生改变,叶片氮含量显著提高,而氮作为合成叶绿素的重要成分,叶片氮含量显著增加使其叶绿素含量合成能力增强,从而导致增温后,杉木幼树叶片光合能力显著增强。此外,本研究发现,尽管增温显著提高杉木光合能力,但对杉木生长指标等无显著影响,可能原因在于土壤增温引起的根际高温加强了杉木的呼吸作用,从而加剧对杉木叶片碳水化合物含量利用消耗过程,最终引起杉木生长特征无显著差异。由于本次实验增温时间较短,对长期增温杉木生长及光合能力的适应机制尚不清楚,今后的研究还应重点关注长期增温对杉木生长及光合特性的影响。
