连续触点运动中障碍物影响个体运动的认知神经机制 *
2019-04-28白学军张琪涵孙弘进陈怡馨孙世南
白学军 张琪涵 赵 光 孙弘进 陈怡馨 孙世南 章 鹏 宋 璐 杨 宇 原 胜
(1 教育部人文社会科学重点研究基地 天津师范大学心理与行为研究院,天津 300074) (2 辽宁师范大学脑与认知神经科学研究中心,大连 116029) (3 麦克马斯特大学心理神经科学与行为系,加拿大 0NL8S4L8)
1 前言
在日常生活中,双手总是频繁地与外在环境发生交互。当个体伸手触及某一物体时,往往会被环境中的相关障碍物所干扰,影响个体的触点(reaching)运动,如抓取水杯后面的手机。我们在计划或执行这一动作时会相应的调整手的运动轨迹,以避免与障碍物发生碰撞(Vaughan, Rosenbaum,& Meulenbroek, 2001; Jax, Rosenbaum, & Vaughan,2007)。这说明个体对当前运动区间(起始点-目标点)内障碍物的加工会影响个体该区间内的运动轨迹。那么,在连续的触点运动中,仅当前运动区间(N)内有障碍物而其他运动区间(例如,前一个运动区间N-1,后一个运动区间N+1等)内没有障碍物时,个体对障碍物的运动计划是否会对区间N+1和N-1内的运动轨迹也产生影响?
关于运动自由度问题(Bernstein, 1967),有人认为,运动的产生基于一个或多个有关消耗最小的标准,例如Flash和Hogan(1985)的最小急动度理论,或Kashima和Isurugi(1998)的最小代谢能消耗模型等。在连续触点运动中,这种消耗最小化理论认为,由于N-1与N+1区间中没有障碍物,个体会计划或执行最短的运动路径以达到运动轨迹的急动度最小或代谢能消耗最低。
然而,在视知觉领域中,Gibson(1979)认为个体对环境或物体的觉知并不是消极被动的,能直接感受到物体传达出的可供交互的动作信息,即称之为物体的可供性。例如,人行道上的石头提供给人“跨过去”的属性。个体观察物体时能够自动激活与物体使用相关的运动计划(Kourtis,Vandemaele, & Vingerhoets, 2018; Masson, Bub, &Breuer, 2011)。因此,相比于区间N中无障碍物基于连续触点运动中障碍物的可供属性,即使N-1区间内没有障碍物,个体由于感知到了区间N中的障碍物,在区间N-1中便已对障碍物进行了计划,继而在该区间的运动轨迹发生了变化,即障碍物对区间N-1中的运动产生了期望效应。
以往基于动物的实验研究发现,前运动皮层、辅助运动皮层及顶叶皮层等与运动的准备活动有关(Andersen, Snyder, Bradley, & Xing, 1997;Shenoy, Sahani, & Churchland, 2013; Svoboda & Li,2018)。以人为被试的研究也显示,个体的前额叶、前运动皮层、顶叶皮层等区域参与运动计划的加工(Aflalo et al., 2015; Maidan et al., 2018;Pilacinski & Lindner, 2019)。例如,Aflalo等(2015)曾将微电极植入到一名四肢瘫痪病人的后顶叶皮层上,发现该区域的神经元集群不但能够编码运动目标还对运动轨迹响应。Heinrichs-Graham和Wilson(2015)使用脑磁图(Magnetoencephalography, MEG)在手指序列运动范式的基础上考察运动计划的复杂性如何调节运动前后的beta震荡及功能连接。结果发现,运动计划的复杂性影响右侧顶叶及左侧背外侧前额叶的神经活动。Pilacinski和Lindner(2019)比较了不同轨迹(直线、曲线)触点动作的大脑活动差异。发现背侧前运动皮层即能够表征直线的运动轨迹,也参与曲线的触点动作,然而顶上小叶仅与直线的触点动作有关。
已有神经影像研究考察了运动计划的神经基础,但对连续触点运动中障碍物如何影响个体的运动轨迹及其脑机制还不甚明了。为了解决这一问题,我们引入了生态效度较高的功能性近红外光谱脑成像技术(functional near-infrared spectroscopy,fNIRS),为在真实的连续触点运动情境下实施实验提供了保证。根据前人关于运动计划脑机制的探讨,我们将探头布局在额叶-顶叶上,覆盖了背外侧前额叶、前运动皮层、顶上小叶等区域。此外,因考虑到上臂连续的弧形轨迹运动可能会引起运动惯性,我们改编了van der Wel等人(2007)的实验范式,将弧形的运动轨迹改为了W形的运动轨迹。最后,基于来自物体可供性、计划再利用及运动计划神经机制方面的研究证据,我们预测在连续触点运动中,区间N内障碍物对区间N-1(无障碍物)中的运动产生期望效应,对区间N+1(无障碍物)中的运动产生启动效应,且障碍物对个体运动轨迹产生的影响与额叶-顶叶区域的活动有关。
2 方法
2.1 被试
招募身高在160-180 cm之间不经常锻炼的被试23名,均来自天津某高校的本科生与研究生。删除3名信号质量较差的被试,最终实验数据来自11名女生和9名男生。平均年龄22±3岁,身高为169.05±6.08厘米,平均臂长为68.95±4.00厘米。经《爱丁堡利手量表》评估均为右利手(Oldfield,1971)。视力或矫正视力均在1.0以上;无色盲;在手臂运动的相关脑区上无主观或客观症状;均无心血管、肺肾、神经、精神等方面的疾病;实验前没有进食含咖啡因的食物或饮料。正式实验开始前被试均签订《知情同意书》,实验结束后得到一定的现金报酬。
2.2 实验设备
运动捕捉仪。采用北京度量科技生产的Nokov光学三维运动捕捉系统(Mars 4H, NoKov Corporation,Beijing, China),来追踪被试的运动轨迹。在木棍的顶端和中间分别固定好一个反光标示点,并使用8个高功率的HLED发光器来捕捉木棍(标示点)的运动轨迹及跨越障碍物的情况。该设备的采样率为100 Hz。
功能性近红外光谱脑成像仪。采用岛津公司生产的LABNIRS近红外光谱脑成像仪(LABNIRS/16,Shimadzu Corporation, Kyoto, Japan)。通过三波长(780±5 nm、805±5 nm、830±5 nm)的半导体激光(IED-60825-1标准下的1M级),经修订后的比尔-朗伯(吸收)定律(modified Beer-Lambert law,MBLL)获得监测皮层下血红蛋白的浓度变化。血红蛋白浓度由3个指标组成:氧合血红蛋白浓度(HbO)、脱氧血红蛋白浓度(HbR)、总血红蛋白浓度(HbT)。本研究采样率为55 Hz。Hoshi,Kobayashi和Tamura(2001)发现相比于HbR和HbT,HbO对任务刺激引起的大脑活动变化更为敏感,故本研究着重分析各实验条件中监测皮层内HbO的浓度变化。
2.3 实验设置及流程
本研究对van der Wel等(2007)的实验范式进行了改编。在其实验范式中,桌面上的6个目标点按弧形摆放,且相邻两目标点之间距离相等。被试需要完成一个相继触及目标点的运动。由于这种弧形的连续运动轨迹可能掺杂了上臂运动惯性的影响,因此,我们根据人体的生理限制及运动特征将弧形的运动轨迹改为W形的运动轨迹。
具体而言,实验被试坐在一张桌子前(长122 cm,宽61 cm, 高78 cm),桌面上贴有5个粉色的圆点。这些圆点的直径均为5 cm,厚度为0.4 cm。相邻圆点之间的距离为31.5 cm,下面的2个目标(距桌边缘5 cm)与上面的3个目标所在直线平行(距桌边缘30 cm),形成了W形的运动轨迹(见图1)。
2.2 染色体与微阵列分析 该患儿外周血染色体核型与微阵列分析检测均未发现异常。在充分知情同意后,应用高通量测序方法对其进行疾病相关基因突变分析。
要求被试右手手持一根木棍(高30 cm, 直径2.5 cm, 重125 g),尽量保证右手小指位于木棍低端,木棍处于垂直。被试以跳跃的方式从一个目标位置到达另一个目标位置,跳跃过程中木棍垂直于桌面。
在一些实验试次中,一个障碍物(高20 cm,宽10 cm, 厚0.2 cm)会放置于W形轨迹的两个目标点之间,因此障碍物的放置位置有4个(见图1)。而在其它试次中,障碍物不出现。障碍物是由透明塑料板制成,通过一个木制底座来固定。
研究者要求被试根据节拍器的节奏(间隔850 ms)来移动木棍依次到达轨迹上的目标位置。最右侧的目标位置是个体每次运动的起始位置。将从起始位置向左移动,到达最左侧的目标点后再依次返回到起始位置的运动称为一个往返运动。在每个往返运动中被试需要跳跃8次,即包含了8个由相邻目标点及运动方向构成的运动区间。
正式实验开始前,被试跟着节奏在无障碍物条件下进行练习。待所采集的数据稳定且被试完全理解指导语后开始正式实验。此过程大约需要2-3分钟。
正式实验中的每个试次都要求被试按节奏完成五次往返运动(34 s)和一个20 s的休息,即当被试听到“开始”后移动木棍,第一个节奏卡在由右向左的第二个目标点上,这种往返运动序列执行五次(34 s),听到“休息”后,木棍回到起始位置并休息20 s,便完成了一个试次。每个被试总共需要完成16个试次,其中无障碍物的控制试次和障碍物出现的实验试次均为8个(障碍物在每个位置上出现2次),各试次伪随机出现。每完成8个试次,被试均会获得一个较长时间的休息(1 min)。
2.4 探头布置
使用由16个发射器和14个探测器组成的两个3×5多通道探头架,探头间距为3 cm,共构成44个通道,覆盖额叶-顶叶皮层(见图2)。根据国际10-20系统,Cz定位在通道15与通道35连线的中点上。采用3D定位仪(FASTRAK, Polhemus,Colchester, VT, USA)确定坐标点(Cz、Nz、AL、AR)及探头位置。通过概率配准方法将各个通道位置与MNI坐标配准,获得其与AAL(Automated Anatomical Labeling)、布鲁德曼分区(Brodmann Area, BA)之间的对应关系。
2.5 数据分析
运动捕捉数据。提取木棍上反光标示点的三维位置坐标。并删除掉那些没有记录到反光标示点位置、撞到障碍物或者没有击中目标点的试次(13个试次)。本研究的运动方式是跳跃,故注重分析Z轴上的位移。对个体在不同运动区间中运动高度的相对值(相比于休息),即计算了个体在不同实验条件下各个运动区间中的平均运动高度。并对各实验条件及各运动区间下的平均运动高度进行统计检验,显著性水平设置为p<0.05。此部分的数据分析通过自编Matlab代码实现。
fNIRS数据。处理方法与白学军等(2016)类似。首先,先对所提取的fNIRS信号降采样(55 Hz到9.26 Hz)。再去噪音和漂移,即在Matlab(v2014a)操作环境下使用NIRS-SPM(Near infrared Spectroscopy-Statistical Parametric Mapping)软件包(v4.1)(Ye, Tak, Jang, Jung, & Jang, 2009)。通过小波分析(Jang et al., 2009)和基于血液动力响应函数(Haemodynamic Response Functions,HRF)的低通滤波器排除掉漂移和噪声(例如头动、心率等)。然后,设置参考波。通过一般线性模形(General Linear Model, GLM)评估各个通道上由实验任务诱发的反应响应参考波的程度(beta值),采用precoloring方法调节此过程的时间自相关。根据实验设计,对不同实验条件下获得的beta值进行配对样本t检验,并FDR校正。
运动捕捉数据和fNIRS数据。将运动捕捉数据和fNIRS数据在Matlab平台上做积差相关。本研究的显著性水平均设置为p<0.05。
3 结果
3.1 行为结果
被试在不同障碍物位置上对各运动区间的平均运动高度结果如图3所示。对行为结果进行5(障碍物位置:1, 2, 3, 4, 无)×8(运动区间)的重复测量方差分析。结果显示:障碍物位置的主效应显著,F=62.44,p<0.01,η=0.62;运动区间的主效应显著,F=38.02,p<0.01,η=0.49;障碍物位置与运动区间的交互作用显著,F=264.34,p<0.01,η=0.87。对各运动区间进行条件之间的配对样本t检验,结果可见表1。
结果表明,W形运动轨迹中区间N中的障碍物影响区间N-1与区间N+1中的平均运动高度,且其对区间N+1的作用效果显著高于对区间N-1的作用效果。具体而言:
当障碍物出现在位置1时,区间1和8(N)中有障碍物,区间2(N+1)和7(N-1)中的平均运动高度显著高于无障碍物条件中这些区间内的平均运动高度。对有无障碍物条件在区间2及7上的差异值进行配对样本t检验,发现区间2上的差异值显著高于区间7上的差异值(t=7.03, p<0.001, d=1.39)。
当障碍物出现在位置2时,区间2和7(N)中有障碍物,区间3及8(N+1)和1及6(N-1)中的平均运动高度显著高于无障碍物条件中这些区间内的平均运动高度。对有无障碍物条件在区间3及1和8及6上的差异值进行配对样本t检验,发现区间3和8上的差异值显著高于区间1和6上的差异值,统计结果分别为t=7.19,p<0.001,d=1.54 和 t=7.11,p<0.001,d=1.42。
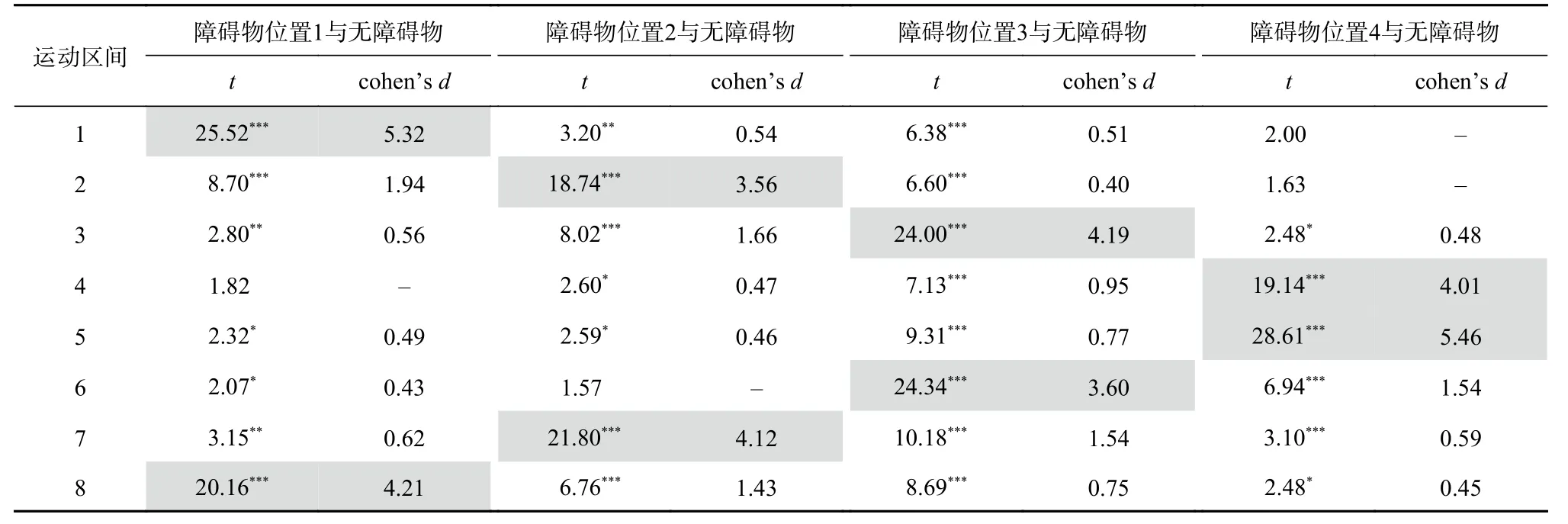
表 1 各运动区间中实验条件间行为指标的比较
当障碍物出现在位置3时,区间3和6(N)中有障碍物,区间4及7(N+1)和2及5(N-1)中的平均运动高度显著高于无障碍物条件中这些区间内的平均运动高度。对有无障碍物条件在区间4及2和7及5上的差异值进行配对样本t检验,发现区间4和7上的差异值显著高于区间2和5上的差异值,统计结果分别为t=5.46,p<0.001,d=1.12 和 t=5.87,p<0.001,d=1.12。
当障碍物出现在位置4时,区间4和5(N)中有障碍物,区间6(N+1)和3(N-1)中的平均运动高度显著高于无障碍物条件中这些区间内的平均运动高度。对有无障碍物条件在区间6和3上的差异值进行配对样本t检验,发现区间6上的差异值显著高于区间3上的差异值(t=6.58, p<0.001,d=1.41)。
3.2 fNIRS结果
对实验条件(有、无障碍物)进行配对样本t检验。结果发现,经FDR校正后,个体在有障碍物与无障碍物条件之间存在显著差异的通道有6、7、25、26、28、29,对应着双侧背外侧前额叶、双侧布罗卡区岛盖部(见表2及图4, 其中AAL表示Anatomical Automatic Labeling, BA表示Brodmann Area)。这说明这两个区域主要参与对W形运动轨迹中障碍物的加工。
3.3 行为指标与fNIRS信号的相关
将被试条件间行为指标及beta值的差异量(有障碍物条件减去无障碍物条件)进行相关计算,发现该行为指标与fNIRS指标在通道26(r=-0.47,p=0.035)和29(r=-0.76, p<0.001)上相关显著,可见图5。这说明W形运动轨迹中障碍物引起的右侧额中回激活强度的变化与平均运动高度的变化之间存在负相关关系,即当障碍物对个体平均运动高度的影响越大时,个体右侧额中回的激活水平越低。
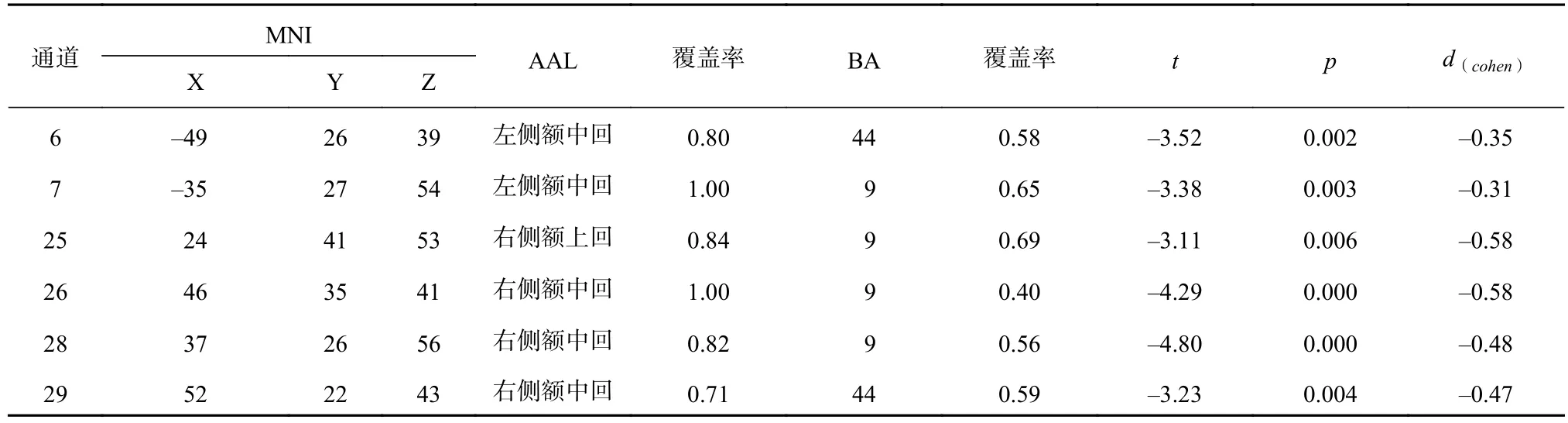
表 2 实验条件间 fNIRS 指标的比较
4 讨论
本研究的行为数据显示,相比于区间N中无障碍物条件,区间N中有障碍物时,区间N-1与区间N+1中的平均运动高度更高。这说明区间N中的障碍物对区间N-1中的运动产生了期望效应,又启动了后面区间N+1中的运动。将这两种效应进行比较后,发现区间N中的障碍物对区间N+1的影响程度显著高于对区间N-1的影响,这说明启动效应的程度要远远高于期望效应。Jax和Rosenbaum(2007)的研究证实了区间N中障碍物对区间N+1中运动的启动效应。van der Wel等人(2007)的实验也在连续触点运动中观察到了期望效应与启动效应。这些研究均说明,个体对连续触点运动中各个区间的运动计划并不是基于消耗最小化原则。区间N中有障碍物时个体在区间N+1与N-1中运动轨迹并不是基于能量消耗或急动度最小产生的。由于区间N中障碍物的可供性(Kourtis et al., 2018),个体在区间N-1中便已对障碍物产生“跨过去”的预期,继而在该区间下的平均运动高度要高于无障碍物时。而在区间N+1中,因对区间N中形成的抽象时空形式的再利用,减少了区间N+1中运动产生的计算消耗(Jax et al., 2003; Rosenbaum et al., 2001),使其运动轨迹尽可能与区间N中的运动轨迹相一致,即出现了启动效应,为后续连续运动的有效控制提供了保证。
此外,脑成像数据显示,双侧背外侧前额叶(BA9)和双侧布罗卡区岛盖部(BA44)在区间N中有障碍物条件下的激活程度显著低于区间N中无障碍物条件。且障碍物对个体平均运动高度的影响越大,右侧这两个区域的激活水平越低。背外侧前额叶参与计划、工作记忆等与执行功能有关的内在加工(Nejati, Salehinejad, & Nitsche, 2018)。例如,Heinrichs-Graham和Wilson(2015)发现了运动计划的复杂性对右侧顶叶及左侧背外侧前额叶活动的调节,证明了背外侧前额叶与个体运动计划之间的密切关系。Kaller等(2013)在伦敦塔任务上也观测到了背外侧前额叶在计划加工中的作用,说明该区域与利用任务规则制定计划有关。可见,背外侧前额叶并不是运动计划所特定的激活区域,它更可能是以完成所有任务均需的执行功能的方式来监控并调节着任务的执行。
布罗卡区岛盖部(BA44)参与动作具体特征的加工(Kilner, 2011)。Kilner(2011)认为运动执行时的动作选择包含两条通路,一条是背侧的运动观察通路,另一条是连接额中回(BA47)到额下回前部的腹侧通路,动作理解需要在这两条通路的交互作用下完成。额下回后部的镜像神经元(猴子的F5c区, 人类的BA44/BA6区)能够编码动作的具体特征,例如连续触点运动中个体运动轨迹的高度或速度等,而对该动作的抽象特征(例如目标或意图)不起作用。这种在更为抽象水平上的加工则是由腹侧通路负责的。结合本研究的实验结果,有障碍物条件下布罗卡区岛盖部的激活强度显著低于无障碍物条件的原因,可能在于这两种条件下个体运动轨迹的具体特征不同。本研究的相关分析也显示,该区域的激活水平随障碍物对个体平均运动高度影响程度的增高而降低。然而,有无障碍物条件间的脑激活差异为什么没有反映到腹侧通路上?我们猜测,这两种条件的抽象特征可能是一致的,其运动目标均为触及目标点,而非跨越障碍物。这一问题还有待进一步的探究。
5 结论
在本实验条件下,可得出结论是:在连续触点运动中,区间N中的障碍物对区间N-1中的运动产生了期望影响,又启动了区间N+1中的运动,且区间N+1中的启动效应远大于区间N-1中的期望效应。背外侧前额叶及布罗卡区岛盖部可能参与了这一加工。
