开垦和长期施肥下青海黑钙土中腐殖质的光谱特征*
2019-04-25何小松刘元元席北斗
张 桐 何小松 李 猛† 刘元元 席北斗 张 慧 李 丹
(1 中国环境科学研究院环境基准与风险评估国家重点实验室,北京 100012)(2 重庆大学城市建设与环境工程学院,重庆 400044)
腐殖质是土壤中的重要组成部分,占土壤有机质比重的50%~70%,主要来源于植物残体或(微)生物体降解和土壤生物化学合成等过程[1],常起到维持土壤碳平衡、调控土壤肥力、调节土壤理化性质的作用[2]。腐殖质并非单一的有机化合物,而是一系列在组成、结构及性质上既有共性又有差别的有机分子结合而成的复杂“生物超分子”结构,含有丰富的功能基团和有机组分[3]。应用紫外—可见光谱、傅里叶变换红外光谱和三维荧光光谱分析技术对土壤腐殖质进行综合表征,可以鉴定其中的有机组分和官能团组成,并解析出腐殖质的性质、来源与功能[4-6]。
土地利用方式的变化和施肥策略常是影响腐殖质结构、性质和功能发生动态变化的关键因素[7-8]。有研究表明:土地利用的变化可能导致土壤腐殖质在来源、结构和性质各层面上均发生显著性改变[9]。而施肥则因其直接改变土壤营养供给和基本理化性质,如pH、阳离子交换量,可能对腐殖质的组成与性质产生重要影响[10]。但目前在开垦和施肥对腐殖质影响的研究上,主要关注于对腐殖质含量的影响,而对腐殖质分子结构和性质影响的研究较少。
青藏高原分布有大面积的高原或高山草甸,在土壤中长期累积了大量的腐殖质。由于农业活动,部分草甸被开垦为农田,并长期施入了化肥或有机肥。在青藏高原高寒环境下,土壤的生态环境较为脆弱,土壤腐殖质的生物化学行为可能对环境变化较为敏感。而目前,土壤腐殖质对开垦与长期施肥的响应尚不明确,因此也对优化土地利用的管理和保持土壤肥力构成了障碍[11]。本试验选取青海达板山的南麓谷地作为研究区,采取表层土样,应用红外、紫外—可见、三维荧光光谱技术表征土壤腐殖质,以研究开垦与长期施肥对青海黑钙土中腐殖质光谱特征的影响。
1 材料与方法
1.1 研究区概况与样品采集
研究区地处青藏高原寒温湿润气候带,海拔为2 940~2 980 m,年平均气温为0.3oC,常年降雨量为530~560 mm,无绝对无霜期。研究区内分布有大片油菜地及草甸,土壤类型为黑钙土。
在研究区选取未开垦草甸(M)和不施肥(NF)、施肥13a(F13a)和26a(F26a)的油菜样地各3块样地。其中,油菜地长期种植北方白菜型小油菜(Brassica campestrisL.),种植密度为105万株·hm-2,其作物生长期为110~120 d,本种植季播种于2017年4月中旬,品种为“青杂五号”。未开垦草甸植被主要为蒿草属(Kovresia)。其中,每年分6次向油菜地施入无机复合肥(总计施氮 160~270 kg·hm-2·a-1、施磷80~120 kg·hm-2·a-1、施钾135~225 kg·hm-2·a-1)和牦牛粪便堆肥(9 000 kg·hm-2·a-1)。其中,牦牛粪便堆肥中含碳量为325 g·kg-1、含氮量为19 g·kg-1、含磷量为10.5 g·kg-1。在种植季进行2次漫灌。草甸无施肥和灌溉管理。于10月上旬油菜收割后,在各样地采取混合表层土样(0~20 cm),样品低温保存并迅速运回实验室。在去除草根、砂粒后冷冻干燥,研磨过2 mm和0.15 mm尼龙筛,用于土壤理化性质测定和有机质提取。
1.2 土壤理化性质测定
土壤pH采用电极法测定,有机质采用重铬酸钾外热法测定,阳离子交换量(CEC)采用乙酸钠法测定,碳酸钙含量采用气量法测定[12]。土壤粒度采用激光粒度仪(LS 13 320,Beckman Coulter,美国)测定,全氮含量采用元素分析仪(Elementar,德国)测定。
1.3 土壤腐殖质提取
土壤中胡敏酸和富里酸的提取参考国际腐殖质协会推荐的腐殖质提取方法[13]。得到的胡敏酸粗品用超纯水洗涤3~5次,以去除Cl-(以AgNO3溶液滴定法检查),离心后冻干[14]。经纯化后的富里酸同样进行冻干,收集备用。
1.4 腐殖质的光谱表征
采用傅里叶变换红外光谱仪(Tensor II,德国布鲁克),以KBr压片法测定腐殖质样品的红外光谱。扫描范围为4 000~400 cm-1,分辨率为2 cm-1,扫描次数为64次。应用OPUS V7.5软件(德国布鲁克)对测得的红外光谱谱图进行平滑与基线校正后,分析特征吸收峰的峰高与位移等信息,对特征吸收峰与官能团的鉴定参考相关文献[14-15]。
腐殖质的紫外—可见光谱应用紫外—可见分光光度计(UV-4802,优尼柯公司,美国)测定称取约50 mg胡敏酸或富里酸的干燥样品于50 mL离心管中,加入40 mL 0.1 mol·L-1磷酸缓冲溶液(pH=7),振荡并超声30 min使其完全溶解。胡敏酸或富里酸溶液则以总有机碳(TOC)分析仪(Multi N/C 2100S,耶拿公司,德国)测定TOC含量,并据此以超纯水将溶液TOC浓度逐级调节至10 mg·L-1。以0.1 mol·L-1超纯水为空白,应用稀释液的吸光值。测试时,扫描波长范围为:190~700 nm,扫描波长间隔为1 nm。测定后,计算稀释液在254 nm处的吸光值与TOC浓度的比值得到SUVA254,并分别计算其在240 nm与420 nm、250 nm与360 nm处的吸光度比值得到E2/E4与E2/E3。
腐殖质样品的三维荧光光谱以三维荧光光度计(F-7000,Hitachi,日本)测定。测定时,溶液TOC=10 mg·L-1,激发波长(Ex)范围为200~450 nm,发射波长(Em)范围为280~550 nm,扫描速度为2 400 nm·min-1,狭缝宽带为5 nm。得到的三维荧光谱图在去除散射后,应用DOMFluor工具箱(MatLab v2017a,美国)以平行因子分析法确定腐殖质中的有机组分及各组分的相对含量[16],其特征峰的鉴定参考相关文献[17-18]。
1.5 统计分析
应用单因素方差分析(One-way ANOVA)检验不同处理间土壤理化性质、腐殖质光谱特性的显著差异性(SPSS v22,IBM,美国)。应用主成分分析法(PCA)检验不同处理间土壤腐殖质中各组分的差异性(XLSTAT v2014,Addinsoft,美国)。
2 结果与讨论
2.1 开垦与长期施肥对黑钙土基本理化性质的影响
青海草甸与油菜地中黑钙土的pH为8.16~8.77,土壤呈弱碱性(表1)。草甸经开垦后,土壤pH显著下降(P<0.05),而长期施肥的土壤具有最低的pH(P<0.05)。土壤中含有2.7%~5.3%的砂粒、22.4%~26.8%的粉粒、69.2%~72.3%的黏粒,土壤质地可归为黏壤土。各处理土壤中的砂粒组分因受土壤母质等不均一因素的影响而产生差异性分布,可能会对土壤有机质发育产生影响。草甸土中阳离子交换量(CEC)平均为35.46 cmol·kg-1,高于全国土壤10~20 cmol·kg-1的CEC分布范围,这可能与青海黑钙土中含有较高的碳酸钙有关(平均为18.2~53.6 mg·kg-1)。同时,研究区土壤质地黏重,土壤胶体负电荷量大,所能吸附的Ca2+、Mg2+、Na+等阳离子的能力较强[19]。不施肥而进行长期种植油菜后,土壤CEC显著降低。而长期施肥则导致CEC的显著增高(P<0.05),说明:施肥使得土壤保肥与营养供给能力增强(表1)。草甸土壤中含有平均为113.46 g·kg-1的有机质,开垦后土壤中的有机质含量显著降低。但相比不施肥,长期施肥的油菜地土壤中有机质含量较高,可能说明施肥减缓了有机质的降低。同时,长期施肥也使得油菜地土壤中全氮含量达0.54~0.91 g·kg-1,显著高于不施肥油菜地和草甸土壤(P<0.05)。

表1 不同处理下青海黑钙土的基本理化性质Table 1 Basic physic-chemical properties of chernozem relative to treatment
2.2 开垦与长期施肥对黑钙土腐殖质紫外—可见光谱(UV-Vis)特征的影响
紫外—可见光谱(UV-Vis)能有效地反映出腐殖质的特定分子结构,单位浓度的吸收值或特定波长吸收值之间的比值等指标对腐殖质的宏观结构具有指示意义。一般腐殖质在254 nm处的单位有机碳含量的吸光度(SUVA254)可用于表征腐殖质的腐殖化程度,其数值越大代表腐殖化越强[20]。E2/E3是指腐殖质在250 nm与360 nm处吸光度的比值,可用以表征土壤中腐殖质的相对分子量和芳香化程度,其数值越小代表腐殖质的相对分子量和芳香度越大[21]。土壤腐殖质的E2/E4值则是指其在254 nm和436 nm处的吸光度比值,可用以表征腐殖质的来源,E2/E4值较高表示内源贡献较大,较小则表示以外源贡献为主[22]。其中,外源贡献指的是土壤中的腐殖质是由外源物质如枯枝落叶、生物残体的腐殖化过程贡献而来的,而内源贡献是指在土壤中的微生物作用下将原有的有机质转化为腐殖质的过程。
对青海黑钙土中腐殖质进行UV-Vis分析,计算其SUVA254和E2/E3、E2/E4值,结果见表2。相比富里酸(SUVA254=21.78~32.86,E2/E3=3.04~3.66),胡敏酸具有较高的SUVA254(38.59~44.30)和较低的E2/E3值(2.21~3.14)。这可能说明胡敏酸相较富里酸具有更高的腐殖度、较高的相对分子量和芳香性,这一结果与相关研究结果相符[23]。青海黑钙土中腐殖质E2/E4为4.55~7.02,整体处于较低水平,腐殖质应主要是外源如植物残体腐解产生。E2/E4在草甸和不施肥油菜地土壤的胡敏酸与富里酸中无显著性差异(表2)。但在施肥13 a与施肥26 a黑钙土中,胡敏酸的E2/E4显著低于富里酸,表明相比富里酸,施肥使得胡敏酸更依赖外源输入。对比不施肥的油菜地,草甸土壤中胡敏酸具有较高的SUVA254和较低的E2/E4、E2/E3,而富里酸则具有较低的SUVA254。这可能说明草甸经开垦、不施肥处理后,胡敏酸的腐殖化程度、相对分子量和芳香性均降低。而胡敏酸的E2/E4则升高,这可能是由于外源植物残体输入的减少导致了腐殖质内源贡献(如微生物作用)的相对增强。富里酸对草甸开垦的响应与胡敏酸有重要差异。油菜地土壤富里酸较草甸有较高的SUVA254,表明其在油菜种植过程中,腐殖化增强。对比不施肥油菜地,长期施肥(13 a与26 a)下的黑钙土胡敏酸SUVA254较高,E2/E4和E2/E3则较低,表明长期施肥可能导致了胡敏酸腐殖化程度增强、相对分子量和芳香性增大、对外源依赖性提高(表2)。这可能是因为施肥使得土壤中的碳供给多样化,有机质分子结构趋向于更复杂。在对长期施肥的响应上,富里酸表现出与胡敏酸较大的差异。施肥后,富里酸SUVA254未发生显著变化,E2/E4和E2/E3则升高,表明长期施肥可能导致了富里酸的相对分子量和芳香性降低、对内源依赖性提高,其结果可能与微生物活动有关。

表2 土壤腐殖质紫外—可见光谱特性Table 2 UV-Vis spectral properties of soil humus
2.3 开垦与长期施肥对黑钙土腐殖质傅里叶变换-红外光谱(FT-IR)特征的影响
应用傅里叶变换-红外光谱法分析青海黑钙土腐殖质中官能团的特征吸收峰,结果见图1。不同处理下胡敏酸与富里酸FT-IR光谱中的官能团特征峰位置未发生变化。黑钙土中胡敏酸与富里酸的官能团组成类似:在3 410 cm-1波数处的O-H或N-H伸展振动峰,主要来自于碳水化合物、蛋白质或氨基酸[23];在2 922、2 851 cm-1波数处的脂肪族CH2与CH3伸展振动峰,主要来自细胞膜、细胞壁等组织成分[24];在1 718 cm-1的羧基C=O伸展振动峰和在1 630 cm-1波数处的芳基C=C伸展振动峰[8];在1 120~1 240 cm-1波数处的羧基或酯中C-O伸展振动峰[14];在910~1 080 cm-1波数处的多糖C-O或C-C的特征吸收峰,主要来自于植物、微生物残体中的碳水化合物[25]。
不同处理间土壤胡敏酸、富里酸的红外谱图均具有相似性,但多数特征吸收峰的强度有较大差异(图1)。油菜地土壤胡敏酸各特征吸收峰强度相比草甸均降低,这与油菜地土壤有机质的降低一致。长期施肥下,胡敏酸的O-H、多糖类C-C和C-O、脂肪族CH和CH3的吸收峰强度弱于不施肥土壤,可能是由于施肥增加了作物生物量而大量消耗了土壤碳水化合物和脂肪族有机物[26]。同时,长期施肥使得油菜地土壤胡敏酸中羧基C=O(1 718 cm-1)和C-O(1 221 cm-1)伸展振动峰的强度相对不施肥土壤增强,可能是由于施肥提高了土壤中羧酸类物质的生成。相比草甸,不施肥油菜地土壤中富里酸的羧酸类、芳香类、多糖类等的特征吸收峰强度均降低。但长期施肥后,土壤富里酸的羧酸类、芳香类、多糖类等的特征吸收峰强度均增强。

图1 土壤腐殖质的傅里叶变换红外光谱图Fig. 1 FT-IR spectra of soil humus
2.4 开垦与长期施肥对黑钙土腐殖质三维荧光光谱(3D-EEM)特征的影响
应用三维荧光光谱技术分析黑钙土中腐殖质的荧光光谱,结合平行因子分析法识别出青海黑钙土腐殖质中的5个有机质组分(图2),分别为两个类胡敏酸组分(C1,Ex/Em=260/420;C2,Ex/Em=270/480)、两个UVC类富里酸组分(C3,Ex/Em=220/420;C5,Ex/Em=240/380)和一个水溶微生物副产物组分(C4,Ex/Em=300/350)[18]。其中,C3和C5组分的峰位置均与紫外光类腐殖质(Ex/Em=220~260/380~460)相似,表明其可在吸收紫外光后发射出较强的荧光且荧光组分的分子量较低[14]。类胡敏酸组分C2中包含可见光类腐殖质(Ex/Em=370/480),多来源为外源腐殖质,分子量相对较高[14]。相比而言,C2的发射波长(480 nm)高于C1(420 nm),说明C2的分子结构更复杂,相对分子量更高[6]。同时,C4含有类蛋白的组分(类色氨酸,Ex/Em=230/340),可能是来源于一部分微生物产生的酶或生物残体中的蛋白质。整体上,胡敏酸中的主要有机质组分为C1、C2和C5,富里酸中的主要有机质组分为C1和C3,C4在两种腐殖质中相对含量均较低。
比较不同处理间土壤腐殖质的组成差异,发现开垦使得胡敏酸中C2相对增强、C3与C5相对减弱,表明开垦可能使得胡敏酸分子结构趋向复杂,相对分子量升高(图3a)。长期施肥使得胡敏酸中C4相对含量显著降低(图3b),可能与植物残体输入的减少有关。对于富里酸,开垦显著增强了C1、C3、C5这三个组分的峰强(图3c),但C3的相对含量随开垦降低(图3d),表明开垦有利于富里酸中低分子量荧光组分的生成。同时,开垦也显著提高了富里酸中的类胡敏酸组分C2的峰强度和相对含量。

图2 三维荧光光谱分析与平行因子分析获得的土壤腐殖质组分(C1~C5)Fig. 2 Components (C1~C5) of soil humus by means of 3D EEM spectroscopy and PARAFAC
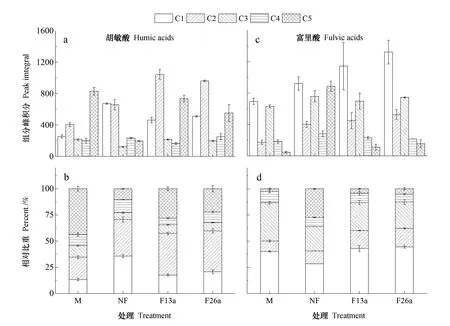
图3 土壤腐殖质中各组分(C1~C5)的相对含量和峰积分Fig. 3 Relative percentages of the components (C1~C5) of the soil humus
应用主成分分析法进一步分析开垦与施肥对腐殖质组成的影响(图4),结果显示:未开垦草甸、不施肥和长期施肥油菜地(13 a与26 a)土壤的胡敏酸与富里酸样品散点均各自团聚,且不同处理间散点相对分散。这可能进一步表明施肥与开垦使得土壤腐殖质组成发生了变化。其中,PC1与PC2分别解释了不同处理下胡敏酸组分差异的66.9%和29.0%。沿PC1,导致样品散点分散的主要因素是C1、C2、C3和C5组分在胡敏酸中相对含量上的差异,表明草甸土壤中胡敏酸中可能含有较多的类富里酸组分(C3与C5),开垦可能导致了这两种组分相对含量的降低。沿PC2,导致其分散的主要因素则是C2和C4在胡敏酸中相对含量的差异。可能表明:长期施肥有利于类胡敏酸C2在土壤胡敏酸中相对含量的增加,但显著降低水溶微生物副产物的相对含量(C4),这可能与施肥后作物残体向土壤的输入减少相关。对于富里酸,PC1与PC2分别解释了不同处理下其组分差异的47.7%和45.6%。沿PC1,草甸土壤样品散点与其他样品散点分散开的主要原因是草甸土壤富里酸中含有较高的类富里酸组分C3和水溶微生物副产物C4,且含有较低的类胡敏酸组分C2,表明开垦可能消耗了富里酸中的类富里酸组分C3和水溶微生物副产物,但有利于生成类胡敏酸组分C2。沿PC2,开垦但不施肥的油菜地样点与其他样点相对分散,可能较低的营养状况下导致了类富里酸组分C1的消耗。

图4 土壤腐殖质组分(C1~C5)的主成分分析的散点图与得分图Fig. 4 Score-plots and loading sectors of the PCA of components (C1~C5) of soil humus
3 结 论
开垦与长期施肥显著影响了黑钙土的基本理化性质、土壤中胡敏酸与富里酸的光谱特性和土壤腐殖质中的分子组成,使土壤有机质含量降低的同时发生土壤酸化现象。开垦但不施肥处理使土壤中胡敏酸的腐殖化程度、相对分子量和芳香性均降低,而开垦后长期施肥则使胡敏酸的上述指标呈现相反的趋势。富里酸在开垦后长期施肥的处理下,其相对分子量和芳香性均降低,而对内源依赖性提高。开垦后不施肥与开垦后长期施肥处理均使土壤中胡敏酸的大分子组分增多,小分子组分降低,并使土壤中富里酸的大分子组成数量和小分子组成数量均增加,且小分子组成的数量增加得更为显著。综上,青藏高原的农垦活动应当谨慎进行。对已开垦后的土壤应当长期施用有机肥以有效减缓有机质的降低。同时,由于施肥使得土壤富里酸对内源依赖性增强,建议在此基础上扩展富里酸的来源。
