不同树龄次生甜槠木荷林的树种组成与空间结构比较研究
2019-04-23袁位高吴初平龚笑飞李大标龚征宇
季 凡,袁位高,吴初平,龚笑飞,李大标,龚征宇
不同树龄次生甜槠木荷林的树种组成与空间结构比较研究
季 凡1,袁位高2,吴初平2,龚笑飞3,李大标3,龚征宇4
(1. 浙江农林大学,浙江 杭州 311300,2. 浙江省林业科学研究院,浙江 杭州 310000,3. 浙江省遂昌县林业局,浙江 遂昌 323300,4. 浙江省遂昌县垵口林业站,浙江 遂昌 323300)
为了进一步揭示次生甜槠木荷林的林分空间结构特征,在浙江省遂昌县垵口村的20 ~ 25年生、35 ~ 40年生、55 ~ 60年生3个次生甜槠木荷林分别设置了面积为50 m×50 m的样地,进行了每木调查和坐标定位。通过样地Ripley聚集度分析和点Ripley聚集度分析,对3个样地中的优势树种及林分整体的空间格局进行对比分析。结果表明:随着树龄的增加,不同年龄的次生甜槠木荷林的上层乔木立木数量逐渐增加,下层乔木立木数量逐渐减少,树种数量相对稳定,但各层树种组成变化明显,其中上层木中甜槠、木荷、青冈3个优势种的比例从20 ~ 25年生时的20.41%增长到55 ~ 60年生时的73.53%;从空间结构看,20 ~ 25年生甜槠木荷林林分呈现聚集分布,35 ~ 40年生甜槠木荷林林分呈现半聚集半均匀分布状态,55 ~ 60年生甜槠木荷林林分呈现均匀分布。
空间结构;物种组成;甜槠木荷林,次生林
林木空间结构是指林木在林地上的分布格局以及它的属性在空间上的排列方式,即林分内树种的不同组成及其空间分布的不同格局。林分空间结构决定了树木之间的竞争势及其空间生态位,在很大程度上决定了林分的稳定性、发展的可能性和经营空间的大小[1]。林分空间分布格局的分析有助于深化林分结构的认识,了解单株木的生长状况,解决营造林中的植株配置和采伐问题。对准确描述种群的空间分布格局、判定林木分布规律以及掌握其过程演化和预测其变化趋势亦具有重要意义[2-3]。近年来对林分空间结构的研究备受关注,林分空间结构理论在森林群落结构研究中应用非常广泛[4]。
关于次生常绿阔叶林空间分布结构的研究,刘韶辉[5]研究次生阔叶林的优势树种甜槠,木荷,青冈的分布格局,指出这3种优势树种总体和个体的空间分布格局均呈聚集分布;李立等[6]在对浙江古田山常绿阔叶林优势树种甜槠和木荷分析出自然环境中大多数物种表现为聚集分布。但是这些研究都是基于一个发育阶段的分析,缺少在不同年龄阶段的次生常绿阔叶林优势树种及林分整体的空间结构特征的研究。因此,本研究对浙江遂昌垵口村不同年龄阶段的次生阔叶林的植被调查,分析比较不同年龄阶段的次生常绿阔叶林的空间结构特征,了解群落空间生长动态的规律,以期为森林抚育经营决策提供理论依据。
1 研究区概况
研究地位于浙江省西南部的遂昌县垵口村,地处28°36′30″ N,119°15′73″ E,海拔730 ~ 814 m,属亚热带季风气候,冬冷夏热,四季分明,雨量充沛,空气湿润,山地垂直气候明显。年平均气温16.8℃,春秋季均温10 ~ 22℃,夏季均温22℃以上,冬季均温10℃。年降水量1 510 mm,年蒸发量850 mm,无霜期172 d。山地黄壤,土层厚80 ~ 120 mm,pH 5.0 ~ 5.5。次生林面积20 hm2,其中大部分为次生甜槠木荷林,本研究的每个不同林龄的次生甜槠木荷林面积约1.0 ~ 1.5 hm2。
2 研究方法
2.1 调查方法
针对3个不同的发育阶段的次生甜槠木荷林进行分析研究,调查时间为2017年10-12月。样地1的林龄为20 ~ 25 a,样地2的为35 ~ 40 a,样地3的为55 ~ 60 a。在相同立地条件设置了3个面积为50 m×50 m的固定样地,共计7 500 m2,每个样地内再设置10 m×10 m的小样方25个。采用全面调查法,调查样地内所有林木特征值,包括林木的相对x,y坐标、树种、胸径、树高、冠幅等,调查工具有围尺、测高器、测距仪、手持罗盘、生长锥等。样地的基本情况见表1。

表1 样地的基本情况
2.2 分析方法
重要值是根据密度、频度和优势度的相对值确定的。物种的重要值越大,其在群落结构中的地位也越重要。因此,可用其表征植物群落物种的结构变化状况[7]。计算公式:重要值=(相对多度+相对频度+相对优势度)/3。
胸高断面积是指树木距1.3 m的位置的断面的面积,可用来表现树木生长过程。计算公式:胸高断面积=(胸径/2)2×π。
采用Ripley’s(d)聚集度分析方法,分析在给定距离尺度、置信度和重复模拟次数条件下的分布格局特殊点分析。Ripley’s(d)分析是可以观测不同距离尺度下林木空间分布格局的方法,用于检验林木空间分布格局。此方法需要对(d)和(d)进行估计,(d)是指从某一随意点起距离以内的其余的期望点数,通过来估算,(d)对分析格局更有用,在随机分布下可使方差保持稳定,其估计公式是:
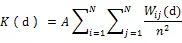
其中,是样地内林木总株数,是距离尺度变量,是样地面积,ij是林木之间距离ij的权重,在Ripley的原始算法中,不考虑样地边界效应,定义:
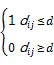
通过ForStat软件进行Ripley’s(d)分析,用Monte-Carlo拟合计算包迹线,用随机模型拟合一组点的坐标值,即每个值对应(d),模拟过程重复进行99次,(d)的最大值和最小值分别为上下包迹线。根据群落实际分别数据,分析不同尺度下的分布,包迹线内为随机分布,包迹线外则偏离随机分布。该程序用于确定显著聚集和显著均匀分布的单株树木。
3 结果与分析
3.1 物种组成
从各样地乔木层主要树种组成、重要值、密度以及胸高断面积(表2)中可以看出,样地1共有21个树种,林分密度为3 144株·hm-2,其中甜槠的重要值为33.98%,密度为1 276株·hm-2,其余如木荷、马尾松和青冈的重要值分别为25.85%,14.82%和10.79%。样地2共有20个树种,林分密度为2 640株·hm-2,甜槠的重要值为43.9%,密度为1 292株·hm-2,木荷、青冈的重要值分别为35.2%和11.93%。样地3共有21个树种,林分密度为1 864株·hm-2,其中甜槠的重要值为36.57%,密度为680株·hm-2,其余如木荷、青冈和马尾松的重要值分别为18.52%,12.39%和2.66%。
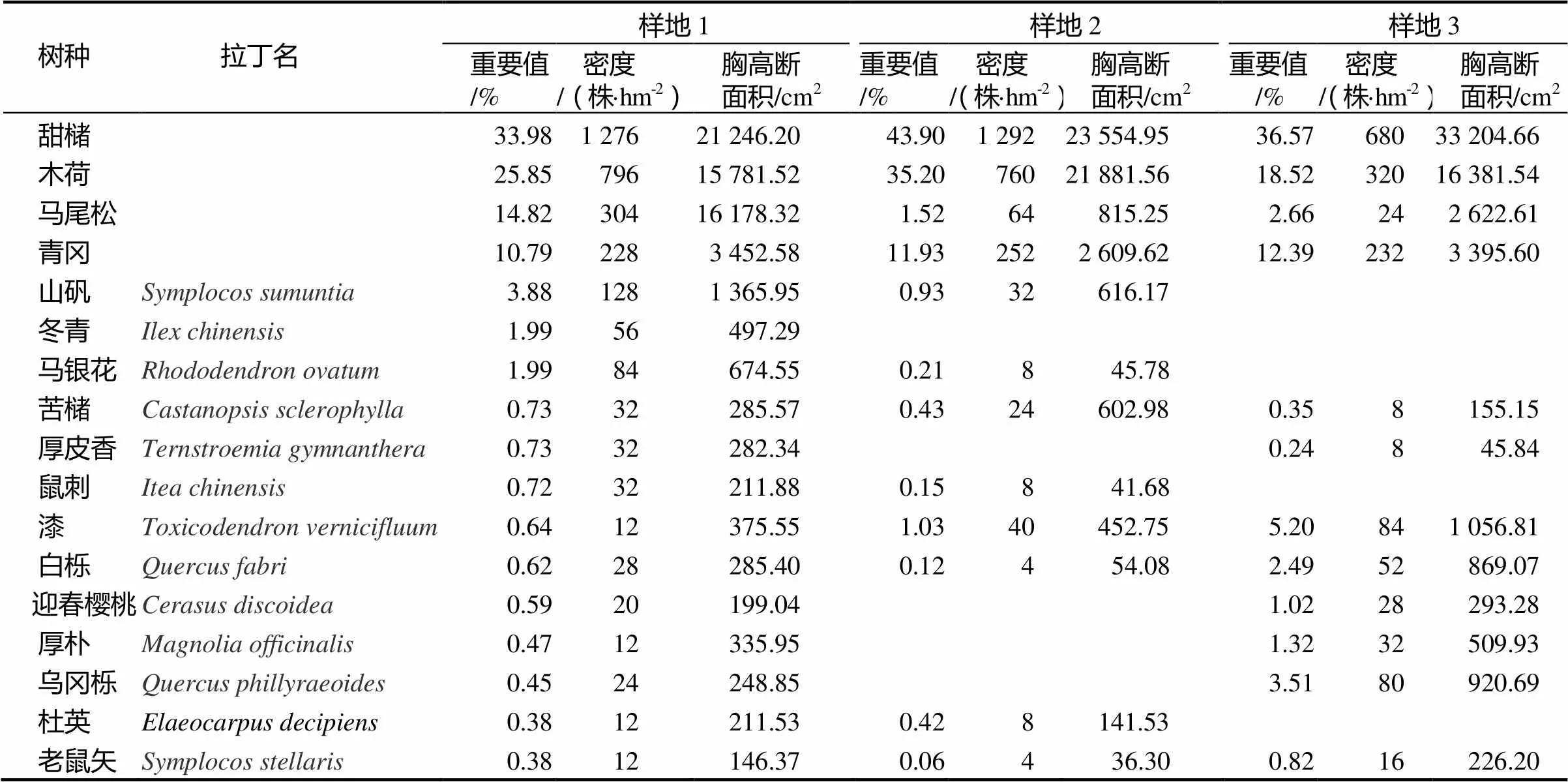
表2 各样地主林层的物种组成
表2 续
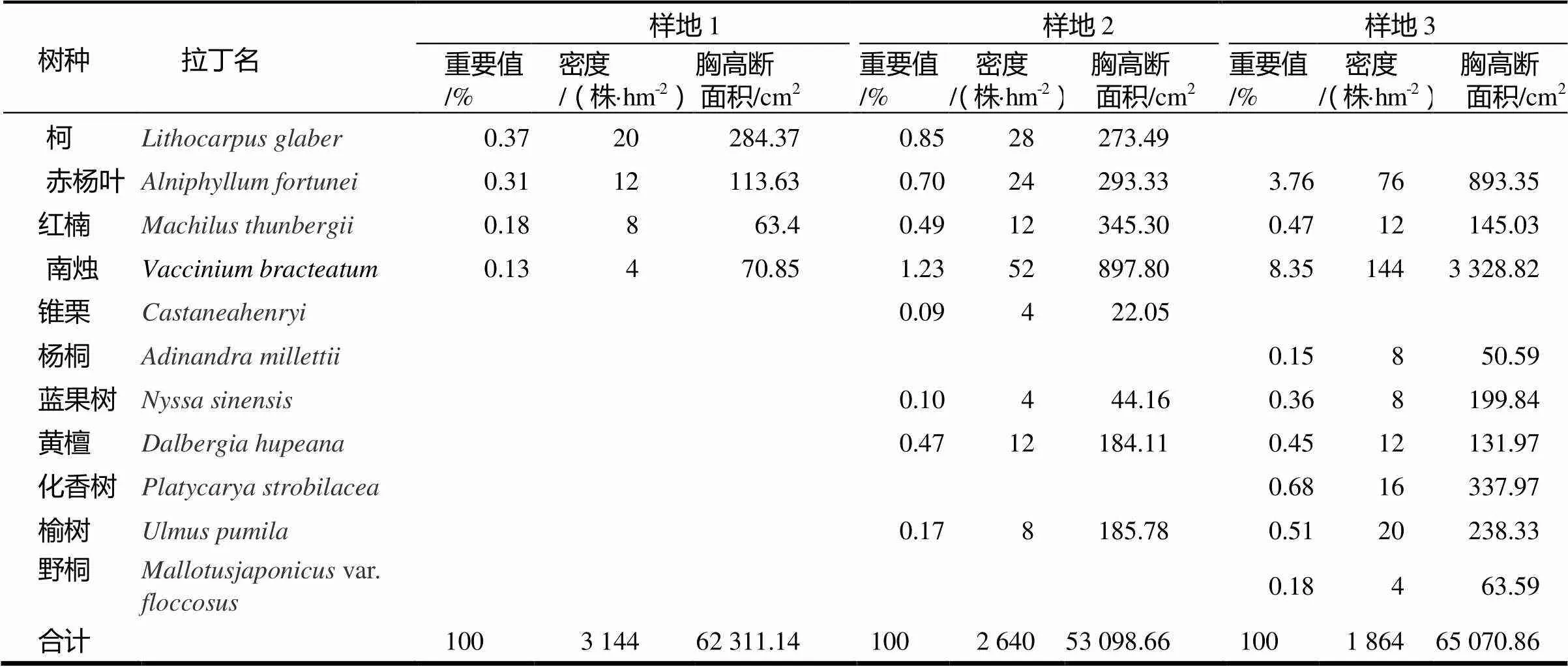
树种 拉丁名样地1样地2样地3 重要值/%密度/(株·hm-2)胸高断面积/cm2重要值/%密度/(株·hm-2)胸高断面积/cm2重要值/%密度/(株·hm-2)胸高断面积/cm2 柯Lithocarpus glaber0.3720284.370.8528 273.49 赤杨叶Alniphyllum fortunei0.3112113.630.7024 293.33 3.7676893.35 红楠Machilus thunbergii0.18863.40.4912 345.30 0.4712145.03 南烛Vaccinium bracteatum0.13470.851.2352 897.80 8.351443 328.82 锥栗Castaneahenryi0.094 22.05 杨桐Adinandra millettii0.15850.59 蓝果树Nyssa sinensis0.104 44.16 0.368199.84 黄檀Dalbergia hupeana0.4712 184.11 0.4512131.97 化香树Platycarya strobilacea0.6816337.97 榆树Ulmus pumila0.178 185.78 0.5120238.33 野桐Mallotusjaponicus var. floccosus0.18463.59 合计1003 14462 311.141002 640 53 098.66 1001 86465 070.86
3.2 结构特征
根据国际林联(IUFRO)的林分垂直分层标准,以林分的优势高为标准把林分划分为3个垂直层:上层为树高≥2/3优势高,下层为树高<1/3优势高,其余为中层;以样地内20株最高的林木树高的平均值为优势高。样地1优势高为15 m,设定乔木层的上层林木≥10 m,5 m≤中层<10 m,下层<5 m;样地2优势高为15.5 m,设定乔木层的上层林木≥11.2 m,5.9 m≤中层<11.2 m,下层<5.9 m;样地3优势高为17 m,设定乔木层的上层林木≥12 m,6.5 m≤中层<12 m,下层<6 m。从各样地群落的分层来看(表3),中层林的密度最大,样地1为2 624株·hm-2,样地2为2 168株·hm-2,样地3为1 536株·hm-2。
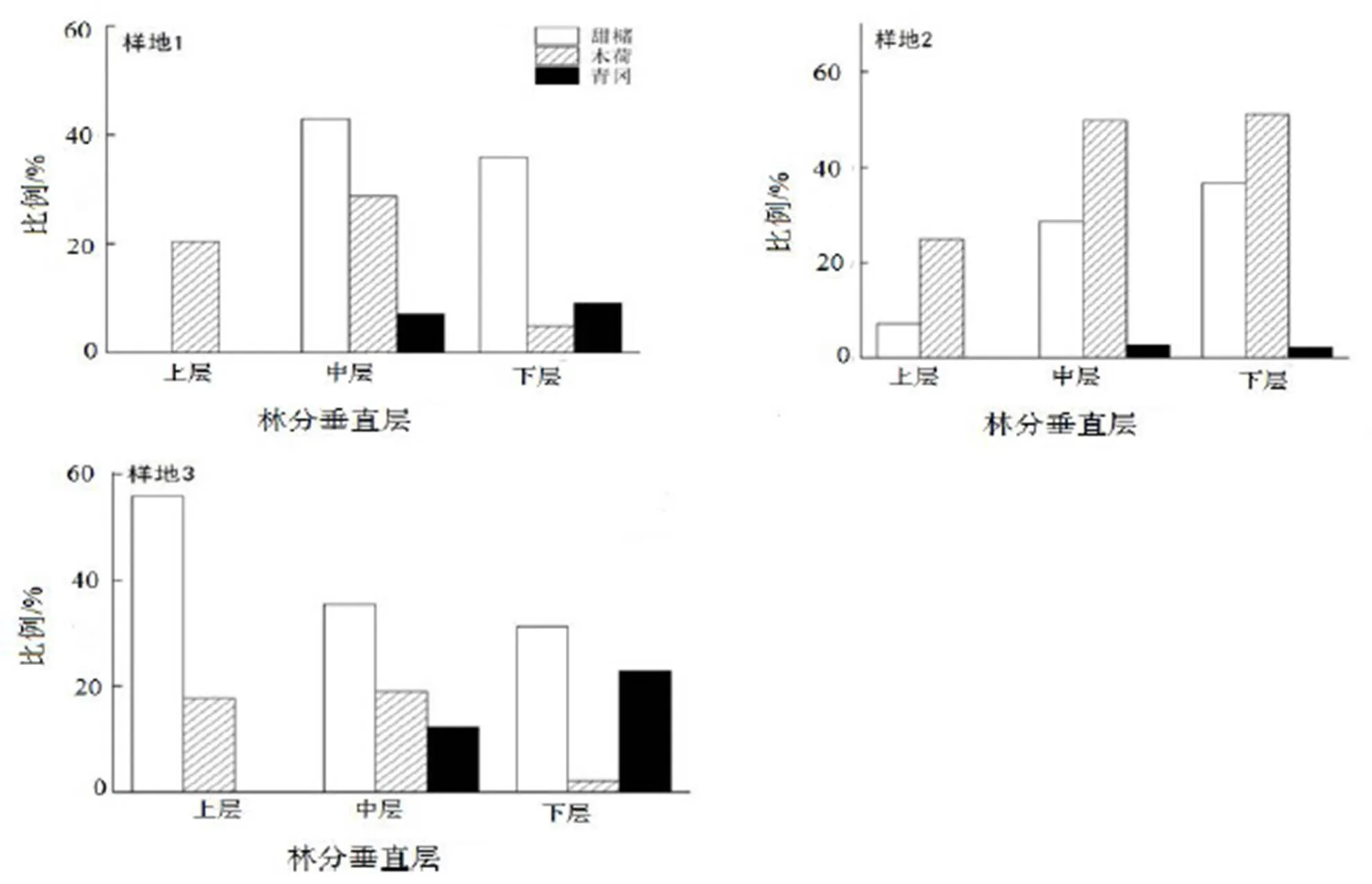
图1 各样地优势树种在群落分层中密度
Figure 1 Density of dominant species at different layers in the communities
上层林木的平均胸径及平均树高最大,样地1的平均胸径21.73 cm,平均树高9.8 m(图1),样地2的平均胸径24.64 cm,平均树高12.26 m,样地3的平均胸径26.35 cm,平均树高14.17 m。中层林木中,甜槠在样地1的平均胸径为22.74 cm,样地2的为29.76 cm,样地3的为35.34 cm。优势树种甜槠在样地2的上层林木密度所占比为7.14%,样地3中增加到55.88%,占据了样地3中上层林木的绝大多数;甜槠、木荷、青冈在林木分层中的密度比例从样地1的20.41%增长到样地3的73.53%,表明在不同年龄阶段的林木中逐渐处于稳定状态。
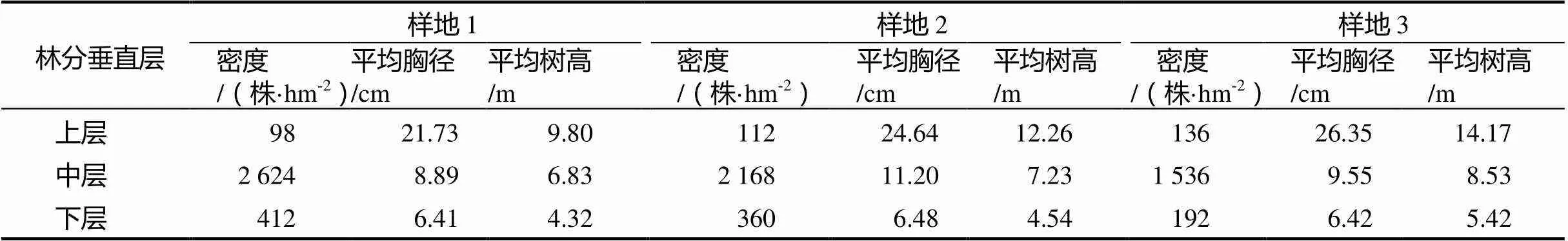
表3 各样地群落分层
3.3 径级结构
图2中可以看出各样地径级分布规律明显,样地1、样地2、样地3均呈现反“J”型分布,其中5≤DBH≤10 cm的个体数最多,分别为401,170,301株,分别占各样地总株数的57.01%,55.56%,63.75%,10≤DBH≤15 cm分别为106,60,166株,分别占各样地总株数的20.08%,19.61%,26.39%。其中,甜槠的径级分布较为连续(图3)。
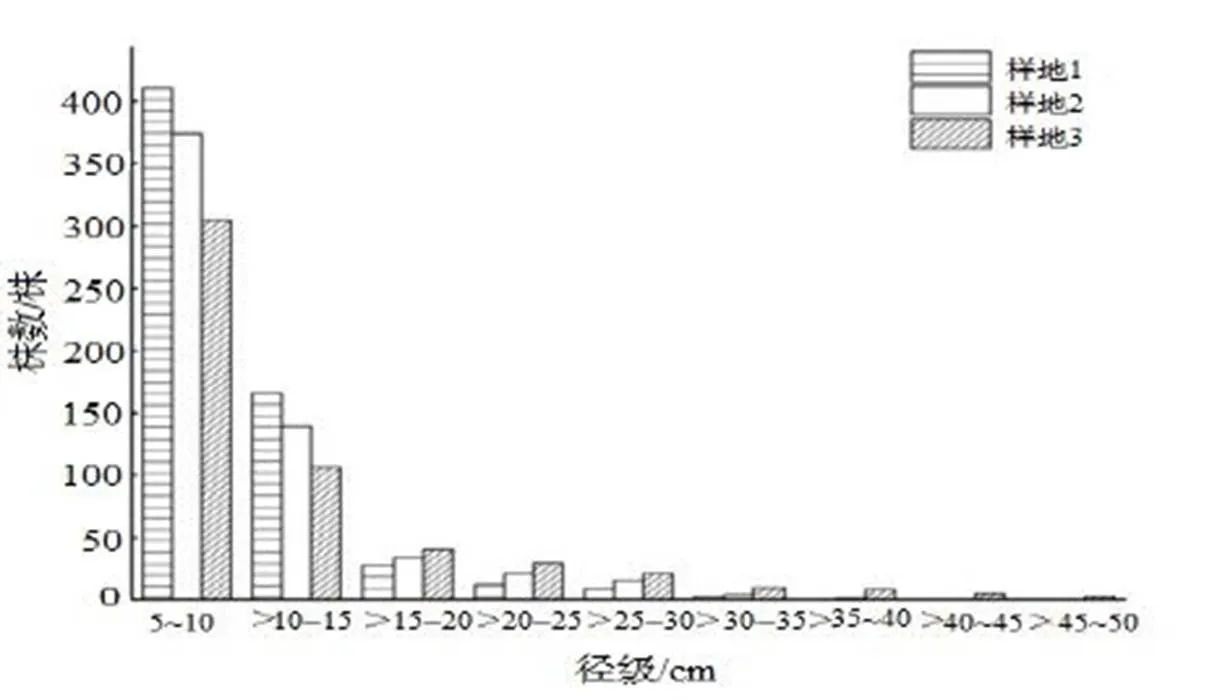
图2 各样地的径级分布图
Figure 2 DBH class distribution of species of different communities
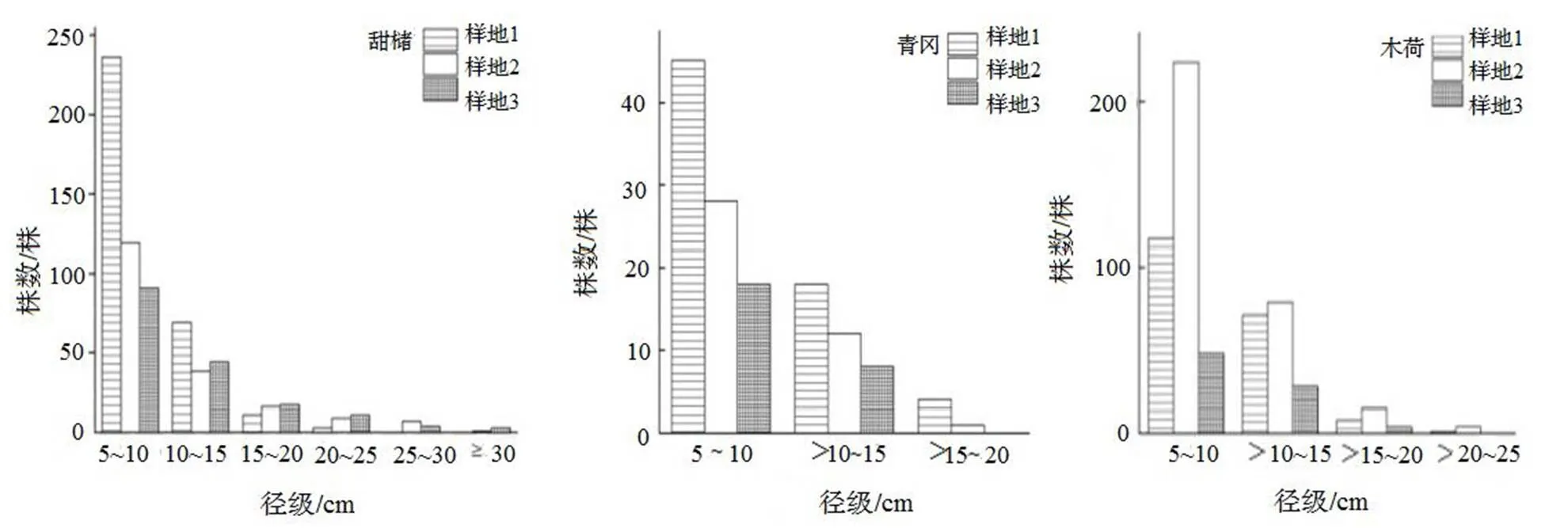
图3 各样地优势树种(甜槠、青冈、木荷)径级分布图
Figure 3 DBH class distribution of dominant species(,,indifferent plots)
3.4 林木空间格局分析
从图4中可以看出,样地1林分整体在所有尺度内基本显示聚集分布;样地2整体在35 ~ 50 m范围内呈现 随机分布,其余尺度均为显著聚集分布;样地3整体在小于5 m的范围内呈现聚集分布,在5.0 ~ 12.5 m呈现随机分布,在12.5 ~ 50.0 m呈现均匀分布。

图4 各样地林分Riley’s K(d)分析图
Figure 4 Analysis of Riley’s()
3.5 优势树种空间聚集度
3个样地共同优势种有甜槠、青冈、木荷,对这3个树种的聚集度进行分析。从图5可以看出,样地1优势树种甜槠、青冈、木荷均呈现聚集分布状态;样地2中,甜槠在0 ~ 20 m尺度呈现聚集分布状态,更大尺度为均匀分布,青冈和木荷呈现聚集分布状态;样地3中,甜槠、青冈、木荷呈现均匀分布状态。

图5 样地优势树木分布图及点 Riley’s K(d)聚集类型图(甜槠、木荷、青冈)
Figure 5 Distribution of dominant trees and aggregation type of Riley’s K (,,)
4 结论与讨论
在研究的20 ~ 25年生、35 ~ 40年生、55 ~ 60年生3个次生甜槠木荷林样地中,甜槠、木荷、青冈的重要值均占优势,为优势树种。随着树龄的增加,不同树龄样地的上层乔木立木数量逐渐增加,下层立木数量逐渐减少;树种数量相对稳定,但各层树种组成变化明显,其中上层木中甜槠、木荷、青冈3个优势种的比例从20 ~ 25年生时的20.41%增长到55 ~ 60年生时的73.53%;3个样地的径级均呈现J型分布,且甜槠的径级分布较为连续。不同树龄林木空间分布格局反映了林木优势树种在生长过程中的动态分布格局的变化。本研究对3个不同树龄阶段林分的空间结构进行研究。结果表明,20 ~ 25年生的在大尺度范围内呈聚集分布,而35 ~ 40年生、55 ~ 60年生的整体空间分布格局也随着尺度的增加呈现为随机或均匀分布。种群从小径级发展到大径级的过程中,分布格局也会随着径级的生长从聚集分布到随机分布的趋势变化,归因于群落中特定机制的发生[8]。
林分空间分布格局是群落树种与环境长期相互适应、相互作用的结果,种内及种间竞争、物种对环境的适应性及其他因素干扰对更新空间的限制等都在一定程度上影响林分的空间结构[9]。
研究表明[10],种群的空间分布格局状态可能与不同树种的生境偏好性以及所处样地地形的复杂性有着密切的关联性。也有研究[11-12]表明甜槠、木荷等常绿阔叶树种有明显的生境偏好。早期种群的空间格局呈现聚集型分布格局是受种子扩散范围以及周围环境相互作用的影响,而使林分树木个体数量减少的原因有群落树种个体径级的生长,种间和种内的竞争压力加剧,密度制约等因素影响下的自然稀疏效应的产生。由于种子扩散的制约,种群在幼年阶段呈现聚集分布,且聚集度高,有助于提高存活率和发挥群体效应,随着种群的生长个体径级的增大,成年阶段聚集强度则开始降低,有助于利用更多的空间资源和环境资源,故种群聚集度的变化可以用种群的生存策略和适应机制来解释[13-14]。
在优化林分空间结构的森林经营措施中,通过次生阔叶林群落优势树种分布变动趋势的研究,根据林分空间聚集度和树种的分布状况,分析树种,确定采伐目标,其目的是使林分空间分布格局趋于随机分布。对于幼龄林和中龄林,可以利用林分空间结构的原理,伐除生长不良、形质不良的林木,促进林下树种的生长与群落恢复,加速林分生长,使不同树种都有各自有利的生态位,形成稳定的森林群落结构,从而维持森林的生物多样性和提高森林生态系统的功能。
[1] 胡艳波,惠刚盈,戚继忠,等. 吉林蛟河天然红松阔叶林的空间结构分析[J]. 林业科学研究,2003,16(5):523-530.
[2] 张会儒,汤孟平,舒清态. 森林生态采伐的理论与实践[M]. 北京:中国林业出版社,2006:47-61.
[3] 叶芳,彭世揆. 种群空间分布理论的发展历史及其现状[J]. 林业资源管理,1997(6):55-58.
[4] 安慧君,惠刚盈,郑小贤,等. 不同发育阶段阔叶红松林空间结构的初步研究[J]. 内蒙古大学学报:自然科学版,2005,36(6):714-718.
[5] 刘韶辉. 湖南会同亚热带次生阔叶林群落特征及种间关系研究[D]. 长沙:中南林业科技大学,2011:80-85.
[6] 李立,陈建华,任海保,等. 古田山常绿阔叶林优势树种甜储和木荷的空间格局分析[J]. 植物生态学报,2010,34(3):241-252.
[7] 贾秀红,郑小贤. 长白山过伐林区云冷杉针阔混交林空间结构分析[J]. 华中农业大学学报,2006,25(4):436-440.
[8] Gavrikov V,Stoyan D. The use of marked point processes in ecological and environmental forest studies[J]. Environmental and Ecological Statistics,1995,2(4):331-344.
[9] 李盾,黄楠,王强. 天然次生林木空间格局及更新格局[J]. 东北林业大学学报,2004,32(5):4-6.
[10] Getzin S,Wiegand T,Wiegand K,He F L. Heterogeneity influences spatial patterns and demographics in forest stands[J]. Journal of Ecology,2008,96(4):807-820.
[11] 陈婷婷,徐辉,马方舟,等.武夷山亚热带常绿阔叶林物种多样性的尺度效应[J]. 生态与农利环境学报,2016,32(5):750-756.
[12] 仲磊,张扬家豪,卢品,等. 次生常绿阔叶林的群落结构与物种组成:基于浙江乌岩岭9 ha森林动态样地[J]. 生物多样性,2015,23(5):619-629.
[13] 赵洋毅,王克勤,陈奇伯. 西南亚热带典型天然常绿阔叶林的空间结构特征[J]. 西北植物学报,2012,32(1):187-192.
[14] 巩合德,杨国平,鲁志云,等. 哀牢山常绿阔叶林树种多样性及空间分布格局[J]. 生物多样性,2011,19(2):143-150.
Comparisonon Species Composition and Spatial Structure of Different Aged Secondary Forest of-
JI Fan1,YUAN Wei-gao2,WU Chu-ping2,GONG Xiao-fei3,LI Da-biao3,GONG Zheng-yu4
(1. Zhejiang A&F University, Hangzhou 311300,China; 2. Zhejiang Academy of Forestry, Hangzhou 310023, China; 3. Suichang Forestry Burean of Zhejiang, Suichang 323300,China; 4. SuichangAnkou Forestry Station of Zhejiang, Suichang 323300,China)
During October and December of 2017, three sample plots with 50 m × 50 m were set up in 20-25, 35-40 and 55-60-year secondary-forest in Suichang, Zhejiang province. Complete enumeration was made in the sample plot. The results showed that treenumbers of overstory increased and that of understory decreased with increase of forest age. Species number maintained stable while species composition of different storey changed evidently. The proportion of three dominant species of the over story such as,andincreased from 20.41% at 20-25-year forest to 73.53% at 55-60-year forest. The analysis demonstrated that 20-25-year stand was aggregated distribution, 35-40-yearsemi-aggregated one, and 55-60 year uniform distribution.
spatial structure; species composition;-forest; secondary forest
10.3969/j.issn.1001-3776.2019.01.002
S792
A
1001-3776(2019)01-0009-07
2018-08-10;
2018-12-29
浙江省重大科技计划(2015C2016)
季凡,研究生硕士,从事多功能森林经营研究;E-mail:nlznjf@163.com。
袁位高,二级研究员,博士,从事森林生态经营与培育研究;E-mail:zfaywg@126.com。
