弓背青鳉的胚胎发育及自发荧光观察
2019-04-13张海瑞王中铎黄顺楷郭昱嵩董忠典刘楚吾
张海瑞,王中铎,黄顺楷,汪 淳,郭昱嵩,董忠典,刘楚吾
(广东海洋大学水产学院 / 南海水产经济动物增养殖广东普通高校重点实验室,广东 湛江 524025)
青鳉属鱼类在我国分布广泛,有记录为中华青鳉 (Oryzias sinensis)、小青鳉(O.minutillus)、弓背青鳉(O.curvinotus)、鳍斑青鳉(O.pectoralis)[1],而日本青鳉(O.latipes)可能是中华青鳉的误记[2-3]。青鳉属鱼类体型小,饲养方便,繁殖力强且周期短,胚胎透明,是研究发育生物学、环境毒理学的重要模式生物[4-5]。其对盐度和温度适应性广[6],可耐受低溶氧,生命力强,疾病少,可通过控制温度和光照周期使其全年产卵[7],且卵大透明,在显微注射和细胞移植等方面具有独特优势,是进行基因功能研究的理想材料[8]。由于青鳉属鱼类的早期阶段对污染物最为敏感,被广泛应用于检测环境内分泌干扰物、致癌污染物等[9-10],是环境毒理学研究的重要模型动物。海水青鳉(O.melastigma)被认为是典型的胚胎毒性分析种类[10],日本青鳉已被国际标准化组织推荐为毒性试验的标准用鱼之一[11]。
弓背青鳉隶属于辐鳍亚纲(Actinopterygii)颌针鱼目(Beloniformes)怪颌鳉科(Adrianichthyidae)青鳉属(Oryzias),在我国广东沿海分布广泛,其性别分化明显,发育早期既可用少量鳍条DNA通过性别特异性分子标记快速、准确地鉴定遗传性别[12],而成年鱼的性别可结合观察臀鳍的形状准确区分。本课题组通过采样调查发现,弓背青鳉栖息水域随潮汐的涨落盐度波动较大,其对盐度波动的适应力强,有用于海水胚胎毒性分析的潜力。我国应用弓背青鳉胚胎进行毒性实验及转基因研究的工作基本鲜见报道,缺乏胚胎发育的基础资料。笔者对弓背青鳉的发育过程进行连续观察、记录,以掌握胚胎各个时期的发育特点及胚胎发育过程中自发荧光的开始时期以及位置,为进一步研究提供基础资料。
1 材料与方法
1.1 实验鱼
用手抄网采集广东省廉江市高桥红树林区域的野生弓背青鳉,于实验室驯养繁育,获得F2代。养殖用水为海水晶配制的人工海水,盐度为12,水温控制在26 ℃;光暗周期比为14 h∶10 h;每日定时喂食卤虫幼体2次。
1.2 鱼卵采集
实验前1 d将1龄的F2代雌、雄亲鱼分开饲养,次日凌晨以雌雄比3∶2混养,约30 min后产卵受精。卵初排出时,卵与卵之间通过长丝相连,成簇地堆积在雌鱼泄殖腔附近完成受精。在自然条件下,受精后数小时,卵黏附在水草上。为观察完整的受精卵发育过程,受精完成后,取雌鱼受精卵,去除卵间长丝。
1.3 胚胎发育观察
取分离的受精卵,置于正置荧光显微镜下,连续观察其发育过程。发育前期,每5 ~ 10 min观察1次,发育至囊胚期后,每1 h观察1次,器官形成期后每5 h观察1次,对各时期典型的特征拍照记录,参考Iwamatsu对日本青鳉胚胎发育时期的特征的描述[13]划分弓背青鳉鱼胚胎发育过程。分别在红光荧光波长590 nm和绿光荧光波长515 nm观察自发荧光现象,并记录。
2 结果
2.1 胚胎发育过程
胚胎发育过程分为受精卵激活期、受精卵胚盘形成期、卵裂期、囊胚期、原肠胚期、神经胚期、器官形成期及孵化出膜期共8个时期。根据各期形态特征的变化,把早期发育过程细分为 35个阶段(图1)。
未受精卵子呈乳白色,不透明,略扁平,卵膜较厚;未受精前,卵膜和卵黄间隙难辨;胚孔位于卵膜动物极,漏斗状;卵黄占的比例较大;大小不一的油滴均匀嵌在卵黄表面,油滴大小有个体差异。
2.1.1 受精激活期 受精后2 ~ 3 min,肉眼观察卵呈半透明状,略带黄色,有一定硬度,卵内结构清晰可见,卵黄大而透明,位于卵中央。光镜下可见嵌在原生质皮层中的皮质小泡开始破裂,位于动物极附近的皮质小泡首先破裂,植物极附近的小泡最晚破裂,其内容物进入卵膜与卵质膜之间,使卵黄周围空间变宽,卵膜与卵质膜之间形成卵周隙。同时,卵膜变薄、变硬,卵黄之间的细胞质开始向动物极流动,在动物极处产生一清晰的细胞质区和一富含卵黄颗粒的植物区。
2.1.2 胚盘形成期 受精后45 min,卵子皮层的原生质向动物极流动,逐渐凸起形成胚盘。在此期间,之前均匀分散于卵黄的油滴开始逐渐相互融合并向植物极迁移。随着时间的推移,油滴开始进一步融合,形成十余个略大的油滴,至该发育阶段结束时,动物半球的大部分油滴已迁移至植物半球,使植物极密度减小而朝上,动物极密度大而朝下。
2.1.3 卵裂期 2细胞期(受精后1 h 20 min)。胚盘逐渐向内缢裂,第 1次卵裂为经裂,卵裂沟穿过胚盘,将胚盘分裂成 2个大小相似近半球形的卵裂球。
4细胞期(受精后1 h 55 min)。经向卵裂为4个大小相同的细胞。
8细胞期(受精后2 h 30 min)。第3次经裂成8个细胞,每行4个细胞。
16细胞期(受精后3 h 15 min)。第4次仍为经裂,分裂球排成4行,每行各4 个细胞,边缘12个细胞较大,内部4个细胞较小。
桑葚胚早期(受精后4 h 15 min)。细胞数约为64 ~ 128,外周细胞略小而扁平。
桑葚胚晚期(受精后5 h 30 min)。细胞变得更小,细胞数约256 ~ 512。
2.1.4 囊胚期 囊胚早期(受精后6 h 45 min)。约为1 000个细胞,细胞更小,形成半圆形的胚盘帽。
囊胚晚期(受精后8 h 20 min)。卵裂球已开始不同步分裂和迁移,导致胚盘帽变低,。
2.1.5 原肠胚期 早前期原肠胚(受精后 10 h)。胚盘帽变扁平,覆盖在卵黄球上,细胞层开始下包。
原肠胚早期(受精后 13 h)。胚盘覆盖卵黄约1/4,胚盘周边的细胞内卷形成胚环,将卵黄囊收紧;背部区域胚层细胞聚集形成胚盾。
原肠胚中期(受精后15 h 40 min)。胚盘覆盖卵黄约 1/2,胚环清晰可见,胚盾面积增加,胚盾中间现一条浅的凹痕,是中枢神经系统原基——神经嵴。
原肠胚晚期(受精后20 h 50 min)。胚盘覆盖约3/4卵黄。
2.1.6 神经胚期 早期神经胚(受精后 23 h 40 min)。植物极仅剩少部分卵黄——卵黄栓未被细胞覆盖;柱状胚体形成,其中一段形成头部。
晚期神经胚(受精后 26 h)。胚体头部两侧出现眼原基,胚孔未关闭前,尾部下方可见透明的Kupffer’s囊。
2.1.7 器官形成期 2肌节期(受精后28 h)。胚体现两个明显的肌节,眼泡后现耳囊原基突起。
4肌节期(受精后33 h)。前中后脑可辨3个区;中后脑两侧现体腔。
6肌节期(受精后38 h)。胚体中央现神经褶;眼泡中可见晶状体,体腔向后扩张。
9肌节期(受精后42 h)。卵黄囊上现少量黑色素细胞;心脏原基出现在脑下部,眼分化出不完全晶体。
12肌节期(受精后46 h)。眼睛内可见完整的晶体轮廓,卵黄囊上现斑块状分布不一的黑色素斑块,头部现少量的棕红色细胞——虹彩细胞。
16肌节期(受精后49 h)。心脏开始微弱跳动,居维氏管在头部和胚体中部形成半循环,虹彩细胞开始扩散到胚体上。
19肌节期(受精后58 h)。心脏的跳动达到70-80次/分,主动脉可见血液流动,Kupffer’s囊完全消失。
24肌节期(受精后68 h)。胚体环绕5/8的卵黄囊,黑色素细胞开始在眼泡中沉积而变暗;尾部开始和卵黄分离。
30肌节期(受精后74 h)。胚体尾部约1/3完全脱离卵黄囊,虹彩细胞扩散到躯干上,黑色素细胞在头部和躯干形成斑状。
35肌节期(发眼期)(受精后97 h)。胚体环绕卵黄囊5/6,眼泡变黑,但晶体透明;耳囊内现2大透明膜突起,该时期也称为发眼期。
鳃血管形成期(受精后112 h)。眼泡可以看到黑色的脉络;血液可以在鳃弓中循环,心跳达90 ~110次/min,躯干出现不规律抖动。
肌节完成期(受精后124 h)。尾部完全脱离卵黄,胚体后部也有部分脱离卵黄囊,并不规律摆动。
脊索空泡化完成期(受精后132 h)。脊索已完全空泡化;眼泡完全黑化,强光下方可辨认晶体。
胸鳍血液循环期(受精后149 h)。胚体完全环绕卵黄囊,眼泡完全黑化;胸鳍经常摆动,出现血液循环;尾鳍现几个黑色素团;尾尖到达眼部。
内脏血管形成期(受精后159 h)。胚体下颌形成,口腔在口凹处开放;血液循环通过内脏,躯体血液可见;尾尖伸展越过眼泡,虹彩细胞从头部到尾端以分散方式分布,头腹部区域现黄色素细胞团,主要为黄色。
心脏发育期(受精后170 h)。尾尖到达耳囊;心脏现明显的心房和心室。
心包腔形成期(受精后8 d)。心脏发育完全,环绕心脏的心包腔突出;尾尖超过耳囊;肠道有一狭窄的内腔。
脾发育期(受精后9 d)。下颌和眼睛活动明显。
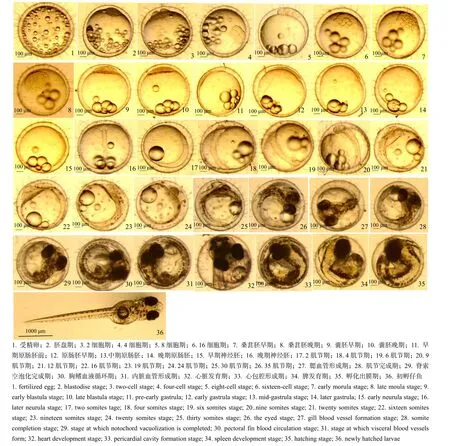
图1 弓背青鳉胚胎发育Fig.1 Embryonic Development of Oryzias curvinotus
2.1.8 孵化出膜期 孵化期(受精后10 d 20 h ~ 12 d)。尾鳍已达胸鳍后,胚体在卵膜内偶有翻转;卵黄越来越小,卵膜变薄,胚体几乎贴在卵膜上,该时期胚体已至出膜期。
仔鱼出膜时,尾部抖动,破卵膜而出,躯体伸展开始后,再完全甩掉卵膜。刚出膜仔鱼体长约3 ~4 mm,躯体有少量色素斑;卵黄囊椭圆形。
2.2 自发荧光
自发荧光主要是色素细胞内物质在激发光下产生的,所以自发荧光位置与色素细胞的出现和分布密切相关,发育至9肌节期时,胚体和卵黄上均出现黑色色素团,块状,位置无规律,于蓝色和绿色激发光下并未自发荧光(图2-1,2,3)。在12肌节期时,头部先出现少量红棕色细胞,此时在荧光激发出现自发荧光,亮度较暗;在 16肌节期时,红棕色细胞团往身体部位迁移,呈树突状和梭状,此时自发荧光较强,发光位置与红棕色细胞团一致(图2-4,5,6);在35肌节期时,已开始迁移至尾部,荧光出现位置仍与红棕色细胞团一致,但在身体上分布位置随机(图2-7,8,9);在孵化期,荧光分布于头部及脊索对应的体节上,呈点状(图2-10,11,12);出膜后,其位置基本与孵化期一致(图2-13,14,15)。
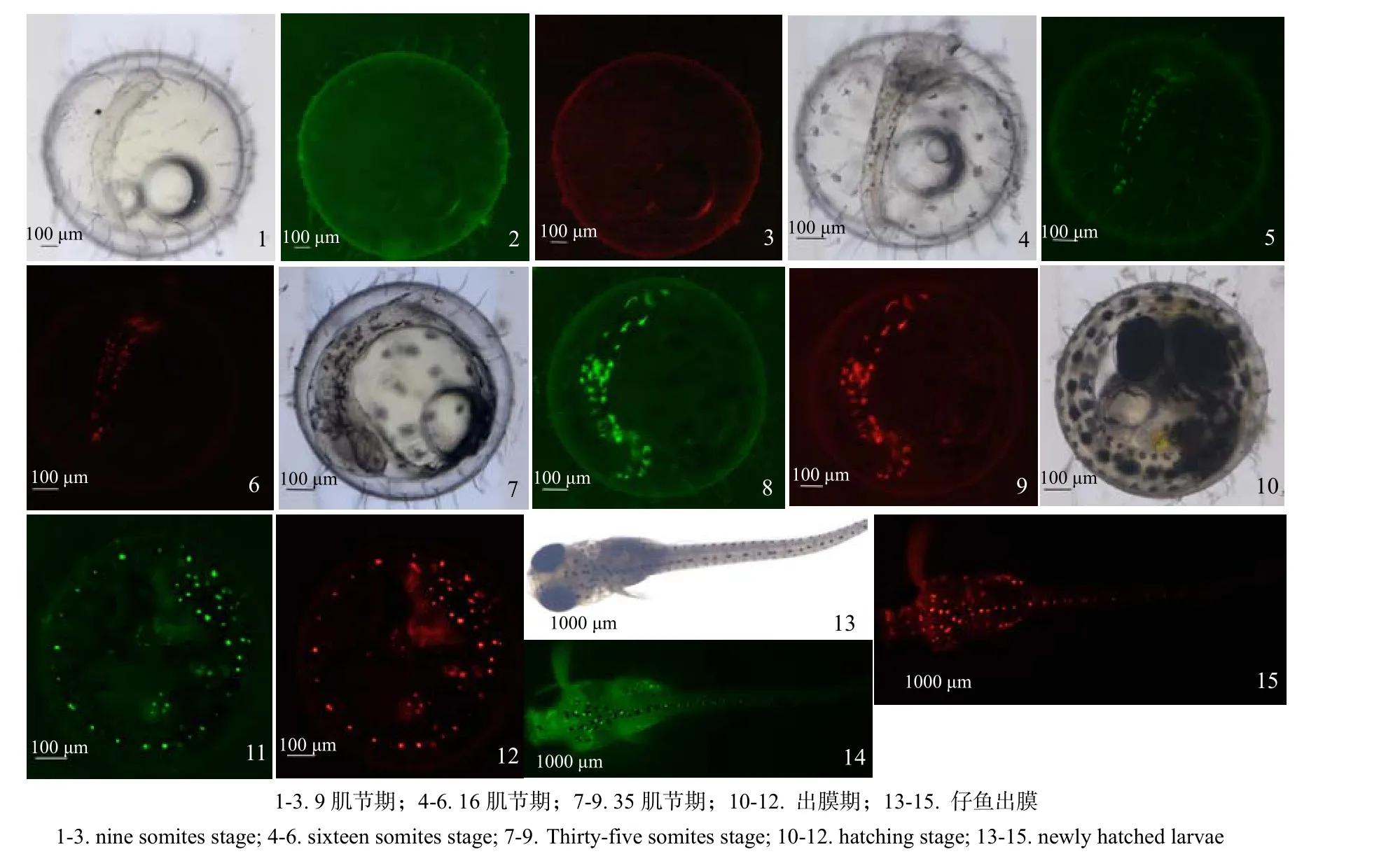
图2 弓背青鳉胚胎发育过程中的自发荧光Fig.2 Spontaneous fluorescence during the development of Oryzias curvinotus embryonic
3 讨论
3.1 受精卵油球
鱼类油球成分主要是脂肪, 密度比水小,因此有油球的卵子一般是浮性卵[14]。然而,弓背青鳉的受精卵与海水青鳉[15]、中华青鳉[16]、日本青鳉[17]等其他青鳉属鱼类一样为多油球沉性卵,其油球随机地镶嵌于卵周细胞质中,受精成功后, 油球逐渐融合。大泷六线鱼(Hexagrammos otakii)[18]、黄姑鱼(Nibea albiflora)[19]、黄斑篮子鱼(Siganus oramin)[20]、棘颊雀鲷(Premnas biaculeatus)[21]和半滑舌蹋(Cynoglossus semilaevis)[22]等硬骨鱼类受精卵的油球融合发生在特定时期。王友红等[15]提出,在多油球的鱼类受精卵中,油球融合时期可作为发育时期的一重要特征。而弓背青鳉卵裂结束后形成数个聚于植物极的大油球的融合及数量变化并无明显规律:有的在神经胚期便融合为一个大油球,有的在器官形成期油球尚未融合,因此,根据油球特征对发育分期并不适用于弓背青鳉。
3.2 Kupffer’s囊
Kupffer’s囊是一个内含单纤毛细胞(Monociliated cells) 的器官[23]。近年来研究证实,斑马鱼(Danio rerio)单纤毛细胞在Kupffer’s囊背腹部的不对称分布会使内液逆时针流动,导致胚胎发育的不对称性,从而启动斑马鱼不对称发育[24],例如胃、心脏和脾脏就是典型的不对称发育器官。然而,越来越多研究发现,并非所有硬骨鱼类均有Kupffer's囊,大鳞鲃(Barbus capito)[25]、普安银鲫(Carassius auratus)[26]、大泷六线鱼[18]等则无Kupffer’s囊,而在黄海大头鳕(Gadus macrocephalus)的胚胎发育中仅 10%个体出现Kupffer’s囊[27]。本研究中,弓背青鳉所有胚体尾部均有Kupffer’s囊,且仅见于尾部位置;其出现于晚期神经胚阶段,即胚孔关闭前。随胚胎的发育,弓背青鳉的Kupffer’s囊会逐渐变大,但位置并不会改变;当发育至尾部将游离的 19体节期,Kupffer’s囊已逐渐消失,而此时器官尚未形成。海水青鳉Kupffer’s囊出现于胚孔关闭前,消失于器官形成的心脏跳动期[13]。因此,Kupffer’s囊对于青鳉不同物种不对称性发育的作用是否存在明显分化还有待进一步研究。
3.3 自发荧光原因
硬骨鱼类色素细胞起源于神经脊细胞[28],主要包括黑色素细胞、黄色素细胞、红色素细胞、虹彩细胞[29-30]。色素细胞呈色方式有2种:一是细胞内含有色素,细胞所呈现的颜色是其所含色素的颜色;二是通过反射特定波长光的方式呈色[31]。从观察结果来看,在只出现黑色素细胞时无自发荧光,至发育后期黑色素细胞位置亦无自发荧光;黄色素细胞出现的时间与自发荧光出现时间不一致;虹彩细胞出现时,自发荧光开始出现,且位置一致。可见自发荧光主要来自于虹彩细胞。研究表明,虹彩细胞的呈色物质主要是与水结合成晶体形式的嘌呤类物质,通过反射特定波长的光表现为白色、蓝色和紫红 3 种色彩结晶体[32]。在曼龙鱼(Trichogaster trichopterus)中,虹彩细胞排列紧密,呈白色或银白色,仅分布在鳞片表层[33]。橘色双冠丽鱼(Amphilophus citrinellus)虹彩色素细胞形状不规则,与其他色素一起表现出不同的颜色[34]。本研究中,观察到的虹彩细胞为棕红色,点状分布于脊索上方,起吸收紫外光[35]、保护神经索的作用。
