硫化氢对镉胁迫下小麦幼苗的生长及生理指标的影响
2019-04-12陈亚华沈振国
张 蓓,郑 翔,黄 维,陈亚华,沈振国,陆 巍
(南京农业大学生命科学学院,江苏南京 210095)
随着现代工业的飞速发展和矿产资源的过度开发,各种污染物不断排放,土壤环境污染问题日益严重,其中重金属污染因其具有富集性、难降解、毒性强的特点而受到广泛关注。我国有接近0.2亿hm2的农田遭受着不同程度的重金属污染,每年因受重金属污染而减产的粮食约有1 000万t,合计损失达200亿元[1]。尚二萍等[2]研究表明,中国粮食主产区耕地土壤重金属污染点位超标率平均为21.49%,污染物以Cd、Ni、Cu、Zn 和Hg 为主,Cd污染比重最高,达到了17.39%,且耕地土壤重金属含量呈增加趋势。
Cd是植物生长的非必需元素,Cd2+被植物根部吸收并被运输至地上部分,在植物体内积累到一定水平后,会造成植物生理生化代谢紊乱,导致植物抗逆性降低,甚至造成植株死亡[3-6]。小麦(TriticumaestivumL.)是我国第二大粮食作物,其产量和质量直接关系国民经济的发展和人民的日常生活。孟自力等[7]研究指出,0.4~1.2 mmol·L-1的Cd2+对小麦种子萌发和幼苗生长有显著抑制作用;不同小麦品种、不同种植区域对镉胁迫的响应存在差异。因此,明确Cd2+胁迫对小麦幼苗生长的影响及外源物质缓解Cd2+毒害的机制对小麦安全生产具有重要作用。
硫化氢(H2S)是一种具有臭鸡蛋气味的气体,已被确定为动物中继一氧化氮(NO)和一氧化碳(CO)之后发现的第三种信号分子,参与哺乳动物的生理调节和疾病反应[8-10],植物体内也能合成少量的H2S,调节植物的生长发育[11]。近年来,许多研究发现外源H2S能够提高植物对重金属胁迫的抵抗能力,如H2S通过增强ATPase活性、减少Al3+的吸收以及提高抗氧化能力来缓解Al对大麦的毒害作用[12];H2S调控AsA-GSH循环,缓解豌豆所受的As毒害[13];H2S通过调节半胱氨酸(Cys)代谢相关基因的表达,参与Cr6+诱导拟南芥中Cys含量增加的过程,并且H2S和Cys共同缓解Cr6+对拟南芥根长的抑制,同时上调PCS1和PCS2表达,增加金属硫蛋白和植物螯合肽的含量,提高拟南芥对Cr6+的耐受性[14]。Mostofa等[15]研究显示,250和500 μmol·L-1的CdCl2处理严重抑制水稻幼苗的生长,而添加100 μmol·L-1的NaHS能够提高抗氧化酶活性,减轻氧化伤害,维持离子平衡,缓解Cd2+对水稻幼苗生长的抑制。此外,H2S预处理可减少胡杨胞质中的Cd,增加Cd在胡杨液泡中的比例,缓解Cd诱导的细胞程序性死亡[16]。
目前,H2S对Cd2+胁迫下的小麦幼苗的生理生化影响和作用机理尚不清楚,尤其是在光合特性方面了解较少。本试验利用硫氢化钠(NaHS)作为H2S的外源供体,水培小麦幼苗至一叶一心期,添加不同浓度的NaHS预处理5 d,再用50 μmol·L-1CdCl2继续处理5 d,然后收苗测定各项生理生化指标,研究外源H2S对Cd2+胁迫下的小麦幼苗生长、光合、叶绿素荧光、Cd含量和抗氧化特性的影响,揭示H2S提高小麦幼苗耐Cd特性的作用机制,为小麦Cd污染的防治提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为江苏省淮南麦区广泛种植的小麦品种苏科麦一号。硫氢化钠(NaHS)为H2S的供体,DL-炔丙基甘氨酸(PAG)为催化H2S合成的L-半胱氨酸脱巯基酶的抑制剂,均购于SIGMA(Sigma-Aldrich,Shanghai),常规化学试剂(分析纯)购于国内化学试剂厂家(索莱宝科技有限公司、国药集团化学试剂有限公司、上海源叶生物科技有限公司等)。
1.2 试验方法
1.2.1 最佳NaHS浓度选择
挑选饱满一致的小麦种子用2.5%(v/v)次氯酸钠(NaClO)溶液消毒30 min,冲洗干净,在28 ℃的培养箱中遮光催芽。发芽后,换用1/2 Hoagland营养液,昼夜温度为26/18 ℃,光暗周期为16/8 h,光照强度为300 μmol·m-2·s-1,每隔1 d更换一次营养液。待小麦幼苗长至一叶一心期,挑选长势一致的小麦幼苗移至塑料杯中培养,随机分为6组,其中4组用含不同浓度NaHS(10、50、100和200 μmol·L-1)的Hoagland营养液预处理5 d,然后用含50 μmol·L-1的CdCl2的Hoagland营养液继续处理5 d;以Hoagland营养液处理5 d后,用含50 μmol·L-1CdCl2的Hoagland营养液处理5 d为Cd对照组;剩余组以Hoagland营养液处理10 d,为空白对照组。每个处理设置3个重复,处理结束后取测定部分生长、生理生化指标和Cd含量。
1.2.2 最佳外源NaHS对小麦活性氧(ROS)和非蛋白巯基(NPTs)含量的影响
由1.2.1确定最佳NaHS浓度后进行以下4组不同的处理:(1)Hoagland处理10 d的空白对照组(Control);(2)NaHS处理5 d+Hoagland处理5 d(NaHS);(3)Hoagland处理5 d+CdCl2处理5 d(Cd);(4)NaHS处理5 d+CdCl2处理5 d(NaHS+Cd),处理结束后取倒二叶进行活性氧(ROS)和非蛋白巯基(NPTs)含量测定。处理时期和方法同1.2.1。
1.2.3 NaHS影响小麦生长的机理研究
为探究NaHS对小麦生长的影响机理,用1.2.1中筛选的最佳NaHS浓度进行8组处理:(1)Hoagland处理10 d为空白对照(Control);(2)NaHS处理5 d+Hoagland处理5 d(NaHS);(3)PAG处理5 d+Hoagland处理5 d(PAG);(4)NaHS+PAG处理5 d+Hoagland处理5 d(NaHS+PAG);(5)Hoagland处理5 d+CdCl2处理5 d(Cd);(6)NaHS处理5 d+CdCl2处理5 d(NaHS+Cd);(7)PAG处理5 d+CdCl2处理5 d(PAG+Cd);(8)NaHS+PAG处理5 d+CdCl2处理5 d(NaHS+PAG+Cd)。PAG的浓度设为1 mmol·L-1,培养方法同1.2.1;处理结束后测定各项指标。
1.3 测定指标与方法
1.3.1 幼苗生长指标与Cd含量的测定
各处理随机挑选15株小麦幼苗,测量幼苗地上部和地下部的长度以及鲜重(FW)。将材料分为新叶、老叶、茎、根,置于105 ℃烘箱中杀青30 min,80 ℃下烘干至恒重(DW);Cd含量的测定参照Zhao[17]的方法,用V(HNO3)∶V(HClO4)=87∶13的混合酸消煮样品,用2.5%稀HNO3定容,用电感耦合等离子体发射光谱仪ICP(Perkin Elmer Optima 2100DV,USA)测定Cd含量。Cd的转运系数定义为地上部Cd浓度与根部Cd浓度比值。
1.3.2 幼苗生理生化指标的测定
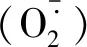
1.4 数据分析
使用SPSS 18.0对数据进行ANOVA分析,采用Duncan test进行多重比较。
2 结果与分析
2.1 不同浓度NaHS预处理对Cd2+胁迫下小麦幼苗生长的影响
与对照相比,不同浓度的NaHS预处理5 d对小麦幼苗的生长无明显影响(表中未列出);而Cd2+胁迫幼苗的生长受到显著抑制(表1),其中幼苗株高降低20.3%,根长降低22.3%,整株鲜重降低25.4%,整株干重降低24.6%。50和100 μmol·L-1的NaHS预处理能够显著缓解Cd2+胁迫小麦幼苗地上部生长的抑制,与Cd2+对照相比,株高分别增加6.7%和5.1%,鲜重分别增加21.9%和17.8%,干重分别增加19.8%和13.6%,而不同浓度的NaHS预处理下根长差异不显著。说明一定浓度的外源H2S预处理能够有效缓解Cd2+对小麦幼苗生长造成的抑制作用,对地上部分的作用效果更明显。
2.2 不同浓度NaHS预处理对Cd2+胁迫下小麦幼苗叶片光合色素和净光合速率的影响
10、50、100 μmol·L-1NaHS预处理5 d时小麦幼苗叶片中的总叶绿素含量、类胡萝卜素含量以及净光合速率(Pn)与空白对照无显著差异,200 μmol·L-1NaHS预处理5 d时小麦叶片的总叶绿素含量和Pn较空白对照显著降低(P<0.05,表中未列出)。如表1所示,与空白对照比较,Cd2+胁迫下幼苗的叶绿素含量、类胡萝卜素含量和Pn均显著降低,分别下降了28.4%、20.7%和21.7%。与单独Cd2+处理比较,光合色素含量和Pn随着NaHS预处理浓度的增加呈现先增后降特点,50 μmol·L-1NaHS预处理下叶绿素含量、类胡萝卜素含量和Pn显著提高,分别提高了12.6%、15.2%和10.8%。说明一定浓度的外源H2S预处理能够有效缓解Cd2+对小麦幼苗叶片光合色素合成的抑制和光合速率的降低,以50 μmol·L-1的NaHS预处理效果最好。
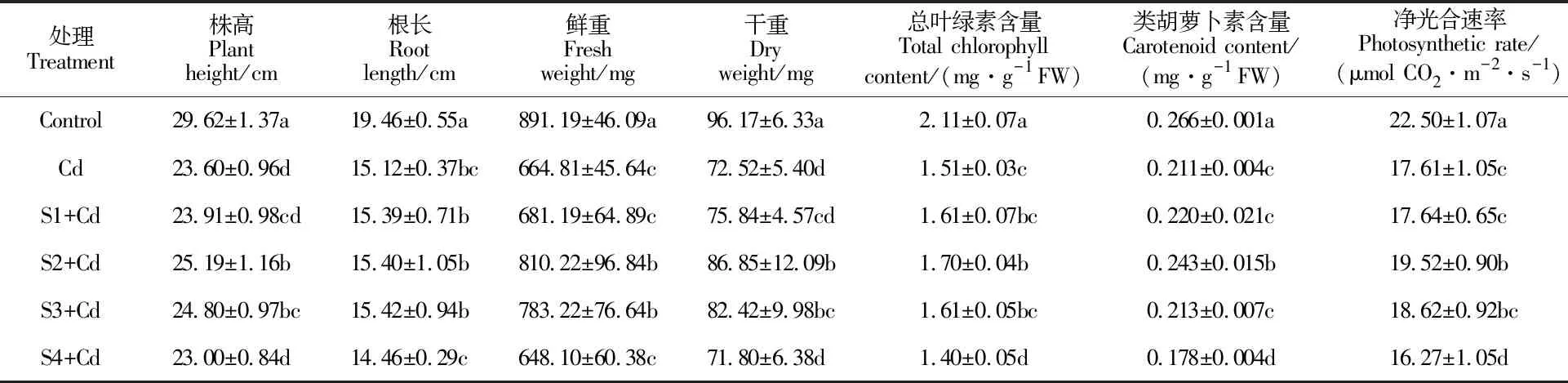
表1 不同浓度的NaHS对Cd2+胁迫下小麦幼苗的生长指标、叶片中光合色素含量和净光合速率的影响Table 1 Effects of different concentration of NaHS on the growth indexes, photosynthetic pigment contents and net photosynthetic rate of wheat seedlings under Cd2+ stress
Control:空白对照组;Cd:Cd对照组;S1+Cd~S4+Cd:不同浓度的NaHS溶液(10、50、100和200 μmol·L-1)预处理再加50 μmol·L-1的CdCl2溶液处理组。同列数据后不同字母表示处理间差异显著(P<0.05)。下同。
Control is the group with no treatment; Cd is the group treated with 50 μmol·L-1cadmium chloride solution; S1+Cd~S4+Cd are the groups pretreated with different concentrations of NaHS solution (10, 50, 100, 200 μmol·L-1) and then treated with 50 μmol·L-1cadmium chloride solution. Different letters following the values in the same column indicate significant differences among the treatments at 0.05 level. The same below.
2.3 不同浓度NaHS对Cd2+胁迫下小麦体内Cd含量和分布的影响
由表2可知,Cd对照小麦幼苗根部和地上部分的Cd积累量分别为2 596.73 μg·g-1DW和546.99 μg·g-1DW。与Cd2+对照相比,NaHS预处理显著降低了老叶和地上部的Cd含量,以50 μmol·L-1的NaHS预处理降低幅度最大(21.6%、9.8%),而茎秆、新叶中的Cd含量与Cd2+对照并无显著差异;50和100 μmol·L-1的NaHS预处理显著增加了小麦根系中的Cd含量,分别提高了24.5%和23.7%。NaHS预处理使小麦中Cd转运系数显著下降,以50 μmol·L-1NaHS预处理的效果最好,降低了27.5%,因此利用这一浓度进行下面的研究。
2.4 H2S对Cd2+胁迫下小麦幼苗叶片中MDA、H2O2和积累的影响
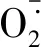

表2 H2S对Cd2+胁迫下小麦茎秆、新叶、老叶、地上部分、根系中Cd含量以及转运系数的影响Table 2 Effects of H2S on Cd concentration in the wheat stem, the new leaf, old leaves, shoot, root and Cd translocation factors under Cd2+ stress
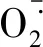
表3 H2S对Cd2+胁迫下小麦叶片中的含量和小麦根中NPTs含量的影响Table 3 Effects of H2S on MDA, H2O2, contents of wheat leaves and NPTs contents both of wheat leaves and roots under Cd2+ stress

图1 H2S对Cd2+胁迫下小麦叶片的DAB和NBT的染色结果

表4 不同处理对Cd2+胁迫下小麦幼苗生长的影响Table 4 Effects of different treatments on the growth of wheat seedlings under Cd2+ stress
2.5 H2S对Cd2+胁迫下小麦NPTs含量的影响
较空白对照,单独NaHS预处理显著增加了小麦叶片和根的NPTs含量,分别增加了23.9%和98.6%;Cd2+胁迫显著增加了根部的NPTs含量;NaHS+Cd与单独Cd2+处理相比,叶片和根的NPTs分别增加了12.5%和49.5%(表3)。
2.6 H2S抑制剂对Cd2+胁迫下小麦幼苗生长的影响
如表4所示,1 mmol·L-1的PAG处理对小麦幼苗被测生长指标均有显著抑制作用。50 μmol·L-1的NaHS预处理后,株高和地上部干重显著增加。在Cd2+存在下,NaHS+PAG预处理与PAG预处理相比,株高和地上部干重分别显著提高了6.7%和29.2%;NaHS对小麦根长和地下部干重影响不显著。说明外源NaHS预处理对Cd2+胁迫小麦的地上部生长有明显正效应。
2.7 H2S抑制剂对Cd2+胁迫下小麦幼苗光合特性的影响
Cd2+胁迫显著降低了小麦叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),NaHS+Cd处理使Pn、Tr和Gs分别提高了16.3%、25.3%和29.6%,差异均显著(表5); PAG预处理使小麦幼苗上述指标显著下降,而NaHS+PAG预处理使Pn和Ci显著增加;在Cd2+胁迫下,NaHS+PAG预处理与PAG
预处理相比,Pn、Gs和Ci分别提高了18.5%、38.9%和24.9%,差异显著。说明外源H2S对Cd2+胁迫下小麦幼苗的光合作用具有明显上调作用。
2.8 H2S抑制剂对Cd2+胁迫下小麦幼苗叶片叶绿素荧光参数的影响
如表6所示,Cd2+胁迫使荧光参数值显著降低,ABS/CSm、TRo/CSm、DIo/CSm和ETo/CSm分别降低了10.6%、10.7%、10.0%和14.8%;除了PAG处理组,各处理下的最大光化学效率Fv/Fm与对照相比无显著差异;Cd2+处理使光系统Ⅱ(PS Ⅱ)的性能指数PIABS显著降低了18.3%;NaHS+Cd与Cd处理相比,ABS/CSm、TRo/CSm、DIo/CSm、ETo/CSm和PIABS分别提高了9.2%、9.0%、10.0%、13.0%和16.4%,差异显著。在Cd2+胁迫下,NaHS+PAG与PAG处理相比,ABS/CSm、TRo/CSm、ETo/CSm和PIABS分别提高了7.4%、8.7%、17.6%和30.8%,差异显著。表明H2S处理显著改善了Cd胁迫下小麦叶片PS Ⅱ的光合性能指数与光能转化效率。

表5 不同处理对Cd2+胁迫下小麦幼苗光合特性的影响Table 5 Effects of different treatments on photosynthetic characteristics of wheat seedlings under Cd2+ stress
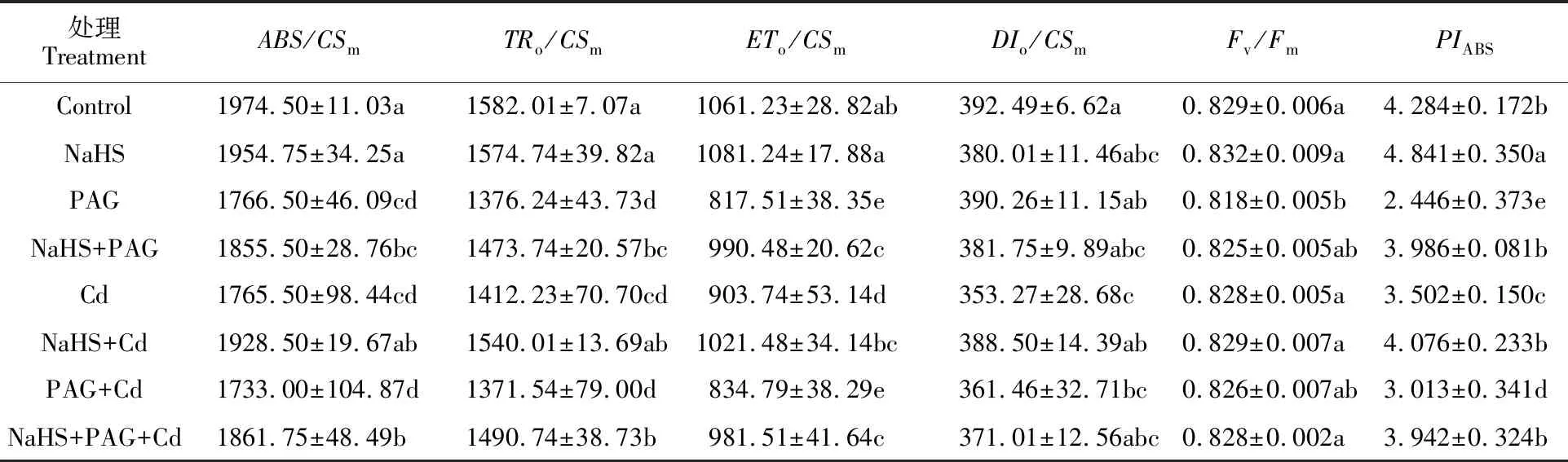
表6 不同处理对Cd2+胁迫下小麦幼苗叶片叶绿素荧光参数的影响Table 6 Effect of different treatments on chlorophyll fluorescence parameters of wheat leaves under Cd2+ stress
3 讨 论
Cd是植物生长发育非必需营养元素之一,浓度过高对植物有很强的毒害作用,严重影响植物的生长发育和代谢[24-26]。在本研究中,Cd2+胁迫显著抑制了小麦幼苗生长。外源低浓度的NaHS预处理能够增加小麦幼苗的株高和生物量,说明外源低浓度的NaHS预处理可以提高小麦幼苗对Cd2+胁迫的耐受能力。Tian等[27]研究发现,镉胁迫抑制谷子叶片光合色素合成并加速光合色素降解,降低植物叶片的光合速率。在本研究中,Cd2+处理显著降低小麦幼苗叶片中光合色素含量,而NaHS预处理后的光合色素含量与单独Cd2+处理相比显著增加,与前人的报道一致[28-29];不同浓度的NaHS对小麦的净光合速率影响与生物量、光合色素含量的变化有相似的趋势。本研究中,Cd2+胁迫使小麦幼苗叶片中Pn、Gs和Ci显著降低,小麦幼苗叶片Pn的降低多数由气孔限制因素引起[30-31];而外源施加H2S能够显著提高Pn、Gs和Tr,说明H2S能够维持小麦叶片较高的光合能力,这可能与NaHS预处理下小麦幼苗叶片中光合色素含量显著增加有关。
Liu等[32]报道,Cd胁迫对棉花叶片的最大光化学效率、量子产额、电子传递速率以及光化学淬灭的抑制与光合作用的减弱密切相关。本研究结果显示,Cd2+显著降低了小麦叶片的ABS/CSm、TRo/CSm、ETo/CSm以及DIo/CSm,而NaHS预处理显著提高了以上参数值,说明H2S提高了Cd2+胁迫下小麦幼苗叶片单位激发态面积吸收的能量、捕获的用于还原QA的能量、还原态QA通过电子传递再氧化的活性以及以热的形式耗散多余的能量,从而加快电子传递速率、保护光合机构。
在正常情况下,Fv/Fm保持在0.8左右,而逆境胁迫会对PS Ⅱ造成损伤或者发生光抑制,导致植物叶片Fv/Fm显著下降[33],但Fv/Fm对Cd2+胁迫不敏感[34]。而PIABS表示PS Ⅱ以吸收光能为基础的光合性能指标,反映原初光化学反应的最大量子产额以及电子传递的量子产额,相比Fv/Fm对逆境更敏感[35-36]。本研究发现,Cd2+胁迫处理能显著降低小麦幼苗叶片的PIABS,而外源H2S显著提高PIABS,说明H2S提高了PS Ⅱ反应中心的氧化还原活性。

NaHS预处理显著增加小麦根部对Cd的吸收积累,而地上部分Cd含量降低,推测H2S的作用是将大量的Cd固定在根中,减少向地上的转运,从而显著降低转运效率。类似的结果还出现在Hg2+胁迫处理的水稻幼苗中,Chen等[38]发现NaHS预处理增加水稻幼苗根中Hg含量,而显著降低地上部分的Hg含量。并且本研究首次发现NaHS预处理使小麦老叶中Cd含量显著降低、使新叶中Cd含量变化不显著。原因可能与NaHS和Cd2+处理时间有关,也可能是H2S会导致Cd在地上各部位的分配和积累产生差异。
植物抵抗重金属毒害的机制有多种,其中一种便是胞内含硫化合物与重金属形成毒性较低的络合物,包括非蛋白巯基化合物(NPTs)和蛋白巯基化合物,其中NPTs又包括半胱氨酸(Cys)、谷胱甘肽(GSH)和植物螯合肽(PCs)[39]。H2S在植物体内能够快速氧化成过硫化物、亚硫酸盐、硫代硫酸盐、硫酸盐等,从而保持较低的H2S浓度,而那些氧化产物通过各种催化途径合成GSH[40],GSH是NPTs的主要组成部分,含有一个活泼的巯基基团,易被氧化,对于清除ROS、与Cd2+形成络合物、促进PCs合成等具有重要作用[41]。本研究发现,H2S可促进根部NPTs的合成来固定根中的Cd2+,减少其向地上部分的转运。
综上所述,外源H2S通过增加小麦叶片和根中NPTs的含量,络合过量的Cd2+,减少叶片中的ROS积累,增加叶绿素含量,保护光合电子传递系统,维持较高的光合能力,促进小麦幼苗生长,提高小麦幼苗对Cd2+胁迫的耐性。
