一株寄生曲霉Q527对黄野螟的感染作用及生物学特性
2019-04-09赵鹏飞常明山吴耀军杨振德
赵鹏飞,常明山,罗 辑 ,吴耀军 ,杨 娟 ,杨振德
(1.广西大学林学院,广西 南宁 530004;2.广西优良用材林资源培育重点实验室/广西林业科学研究院,广西 南宁 530002;3.崇左市森林病虫害防治检疫站,广西 崇左 532200)
【研究意义】黄野螟(Heortiavites soides Moore)隶属于鳞翅目(Lepidoptera)螟蛾科,是土沉香〔Aquilaria sinensis(Lour.)Gilg〕生长过程中危害最严重的食叶性害虫。一龄幼虫和二龄幼虫群居生活,把叶子吃到只剩下薄薄的叶脉,三龄幼虫可啃食整个叶片结构,分散危害,危害面积扩大。黄野螟幼虫从低龄发育到高龄时间较短,一般在发现其危害及准备防治时,其接近或进入高龄期,暴发时将受害区内的土沉香全部吃光,造成树势下降甚至死亡,严重影响土沉香正常生长[1-2]。在土沉香生产中的防治途径主要依靠物理防治和化学防治,物理防治主要是利用杀虫灯和蜜源植物对黄野螟成虫进行诱杀和对各代卵期、幼虫期,或组织人力摘除有虫枝叶[3];化学防治中不同类型杀虫剂的作用方式存在差异,对黄野螟的防效也存在显著差异[4]。然而物理防治人工成本过高,防治成本一般; 化学防治污染环境,害虫容易产生抗药性。由于繁殖迅速、世代重叠、易于爆发成灾[5-7],因此增加了害虫治理成本和难度,治理过程中也会对寄主和周边环境造成影响。可持续资源发展的重要研究领域之一就是生物防治,微生物农药的研究与利用正是现代生物防治的一大特点[8],因此探索新型生物防治措施具有重要现实意义。
【前人研究进展】有关虫生真菌对鳞翅目的影响研究已有一些报道,安建梅等[9]研究表明粉质拟青霉对黄刺蛾幼虫有很高的毒力且具有良好的防治效果,致病死亡率为84%。何劲等[10]研究证实4株虫生真菌对小菜蛾防治中的应用和毒力,结果表明其具有良好的毒杀作用。刘爱英等[11]研究了玫烟色拟青霉和斜链拟青霉对烟草粉螟感染力和致病作用,发现二者具有很好的防控潜力。【本研究切入点】从自然死亡黄野螟尸体上分离发现1株对黄野螟具有高侵染力的菌株,黄野螟被侵染2d后虫体开始出现菌丝体。对该黄野螟寄生菌进行分离鉴定,确定为寄生曲霉Q527(Aspergillus nomiusQ527),曲霉对黄野螟的致死作用和生物学特性至今国内外尚未见报道。【拟解决的关键问题】由于虫生真菌的防治效果容易受到环境因子的影响,只有掌握最有利于虫生真菌的生长条件及产孢特性,才能为有效利用其防治黄野螟危害。为探讨寄生曲霉Q527控制黄野螟危害的可能性,本研究测定其对黄野螟的感染作用,并对寄生曲霉Q527生物学特性进行了初步研究。
1 材料与方法
1.1 试验材料
2018年8月于广西壮族自治区凭祥市东盟林业产业园种植区的土沉香林采集黄野螟幼虫。将采集幼虫放入网袋内,采集500头以上,带回室内于人工气候箱内饲养,喂以新鲜土沉香叶片,饲养温度为26(±1)℃,相对湿度为85%,光周期为16∶8 (L∶D)。
寄生曲霉Q527菌株从自然罹病死亡的黄野螟幼虫体表分离得到,通过形态学和分子生物学方法鉴定为寄生曲霉Q527(Aspergillus nomiusQ527),保存于广西林业科学研究院森林保护实验室。
供试培养基:PDA培养基:马铃薯200 g、葡萄糖20 g、琼脂20 g和蒸馏水1 L;SDAY培养基:蛋白胨10 g、酵母浸出物10 g、葡萄糖40 g、琼脂20 g和蒸馏水1 L;PCA培养基:胰蛋白胨5.0 g、酵母浸粉2.5 g、葡萄糖1.0 g、琼脂15.0 g和蒸馏水1 L;PSA培养基:马铃薯200 g、蔗糖20 g、琼脂20 g和蒸馏水1 L;OA培养基:燕麦片30 g、琼脂20 g和蒸馏水1 L;WA培养基:琼脂20 g和蒸馏水1 L。
1.2 试验方法
1.2.1 毒力测定 菌株在26℃条件下培养15 d,用含0.5%吐温-80的无菌水配制孢子悬浮液,血球板计数浓度为1.0×108个/mL。幼虫采用浸渍法接种[12],将同一批次老熟幼虫浸入20 mL孢子悬浮液2 min后,把幼虫挑入放置土沉香叶片的饲养盒中,每盒20头幼虫。每个处理3次重复,设含0.5%吐温无菌水为对照。每天定时观察记录发病特征及死亡情况,持续观察7 d。
1.2.2 培养基对菌株生长的影响实验 将供试菌株在PDA培养基上活化培养7 d。用直径为5 mm的打孔器在上述PDA培养基中打出菌块,然后分别接入事先倒好的供试培养基。观察并记录生长趋势和形态颜色。
1.2.3 pH对菌株生长的影响实验 分别将菌块接入 pH为 5、6、7、8、9、10、11、12的 PDA培养基中,放置26℃恒温培养箱中,每个处理3次重复。培养5 d,采用十字交叉法测量其菌落直径培养[13];培养20d,镜检并计算各处理平均产孢量。
1.2.4 高温对菌株生长的影响实验 分别将菌块和孢子悬浮液放入无菌离心管内,放置温度为35、40、45、50、55、60、65 ℃的水浴锅内加热10 min后,接入PDA培养基,放置26 ℃恒温培养箱中,每个处理3次重复,直径计算同1.2.3,分生孢子萌发率计算方法参照杜奕君等[14]并略有改进。
1.2.5 不同碳和氮源对菌株生长的影响实验 采用真菌生理培养基[15]分别以葡萄糖、可溶性淀粉、乳糖和麦芽糖为碳源测定菌株对不同碳源的利用;分别以蛋白胨、硝酸钾、硫酸铵和硝酸钙为氮源测定菌株对不同氮源的利用,以不加入碳源和氮源的培养基为对照。将菌块接入准备好的培养基中,放置26 ℃恒温培养箱,每个处理3次重复,直径和产孢量计算同1.2.3。
1.2.6 光照对菌株生长的影响实验 分别将接入菌块的培养皿置于光照、光暗12 h交替和全黑暗的26℃恒温培养箱中,每个处理3次重复。直径和产孢量计算同1.2.3。
数据处理和计算均采用Excel和SPSS 22.0。
2 结果与分析
2.1 寄生曲霉Q527菌株的杀虫活性
用曲霉Q527孢子悬浮液对健康的黄野螟幼虫进行接种,在保湿的无菌环境下培养数天后,从头部和腹部的关节处长出了菌物(图1,封三)。菌物从最初的白色变为明黄色,经分离纯培养,在PDA培养基上逐渐形成浅黄色至淡绿色的菌落。用纯培养菌种接种健康的黄野螟,再次出现大量试虫死亡,死虫经保湿无菌培养数天后又长出形态相似的菌物。将这些菌物再分离纯化培养在PDA培养基上后,得到的菌株形态特征及菌落与孢子特征均与原接种菌株一致。上述结果表明,由科赫氏法则确定寄生曲霉Q527为黄野螟的致死病原菌。

图1 黄野螟幼虫感染过程Fig. 1 Infection process map of Heortiavites soides larvae
不同空间尺度下西南高原山地基本农田质量景观格局分析(内文第120~129页)图版
黄野螟幼虫经菌株Q527接种处理,接种后1 d行动迟缓,食量降低,虫体开始发暗,虫体饱满保持原有形状,接种后2 d逐渐开始死亡,虫体出现变褐、变黑及僵硬的症状,可见白色的菌丝从病发区长出,接种后3 d虫体产生大量菌丝并被其覆盖。接种后4 d虫体逐渐干瘪体积缩小,菌株的颜色转为黄绿色。在室内条件下对黄野螟接种处理后,致死效果明显,菌株的致死率均与对照存在显著差异,接种后2 d内死亡率较低,接种后3 d死亡率升高,为74.1%;随后死亡率逐渐平缓, 接种后7 d幼虫死亡率为83.3%(图2)。

图2 黄野螟幼虫日死亡率Fig. 2 Daily mortality of larval of Heortiavites soides
2.2 培养基对寄生曲霉生长的影响
供试的 6种培养基中菌株Q527在PDA培养基上生长最好,菌丝十分厚实,颜色表现前期为白色,后期为黄绿色。菌落直径达78.18 mm,与其他培养基存在极显著差异;其次是在SDAY和PSA培养基上生长速度较好,菌丝浓密程度一般,菌落直径分别为42.85 mm和41.58 mm;在SDAY培养基上菌丝较为稀疏,在PSA培养基上可以看到明显轮纹;在PCA培养基上生长速度相对一般,但菌丝较为浓密,菌落直径为39.33 mm;在燕麦培养基上生长速度相对较差且轮纹不明显,菌落直径为36.82 mm;在WA培养基不生长。
2.3 pH值对寄生曲霉生长的影响
从图3可以看出,寄生曲霉Q527生长pH范围比较广,在pH5.0~12.0范围内均可生长,有极强的耐碱性,不同pH对该菌的生长影响差异显著。其在pH为8.0生长最快,菌落直径为78.12 mm。培养20 d后观察其产孢量发现,该菌株在pH5.0~12.0之间都可以产孢,在过度的偏酸性条件下,产孢量会受到明显抑制。pH5.0时产孢量最低,每皿分生孢子为4.07×108个;在pH8.0时产孢量最高,每皿分生孢子为2.61×109个;随着pH的继续升高,产孢量也会逐渐变少。表明寄生曲霉Q527适宜生长偏碱性环境中,pH8.0最为适宜。

图3 不同pH值对寄生曲霉的生长和产孢量的影响Fig. 3 Effects of different pH on mycelial growth and sporulation
2.4 高温对寄生曲霉生长的影响
高温对寄生曲霉Q527生长的影响测定结果(图4)表明,其在35~60℃温度下菌株可恢复生长,孢子均可萌发。但菌丝生长受温度变化的影响差异极显著,在35℃菌丝长势最好、边缘整齐,菌落直径为81.18 mm,菌丝在65℃水浴10 min后再次接入培养基中停止生长,表明随温度升高,菌丝生长速度减缓直至死亡。曲霉Q527孢子萌发受温度变化明显,菌株的孢子萌发率均随温度的升高而减小。分生孢子在65℃时停止萌发,在35 ℃时孢子萌发率最高,为98.13%。

图4 高温对菌丝的生长和孢子萌发的影响Fig. 4 Effects of high temperature on mycelial growth and spore germination
2.5 碳源对寄生曲霉生长的影响
从图5可以看出,菌株Q527在测试的5种碳源培养基上均可生长,但对各碳源利用程度不同。从菌落生长速度看,发现其对乳糖具有很好的利用率,在含乳糖的培养基上,菌丝生长速率最快,菌落直径为37.97 mm。碳源对菌丝生长速度和产孢量的影响没有明显相关性,不同碳源对病原菌产孢影响也不同,最适合产孢的为含可溶性淀粉的培养基,培养20 d平均产孢量为2.03×109个/皿;含葡萄糖、乳糖和麦芽糖的碳源培养基均可以产孢。不同碳源对菌株Q527影响差别较大,在不含碳源的培养基上生长速度和产孢量最低,说明碳源可以促进菌株Q527生长和产孢。
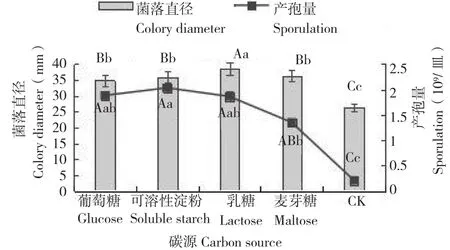
图5 不同碳源对寄生曲霉的生长和产孢量的影响Fig. 5 Effects of different carbon sources on mycelial growth and sporulation
2.6 氮源对寄生曲霉生长的影响
从图6可以看出,不同氮源培养基对菌株Q527菌丝生长影响差别较大,蛋白胨是菌丝生长最快及产孢量最多的氮源,培养7 d菌落直径达到51.09 mm;培养20 d产孢量达2.96×109个/皿;在含氮培养基中,硝酸钙培养基的产孢量最低,培养20 d产孢量为0.74×109个/皿,表明硝酸钙作为氮源不适合菌株Q527产孢;硝酸钾和硫酸铵作为氮源和空白对照比较,发现其抑制菌丝生长。在不含氮源的培养基上产孢量最低,说明含有氮源可以促进菌株Q527产孢。

图6 不同氮源对寄生曲霉生长和产孢量的影响Fig. 6 Effects of different nitrogen sources on mycelial growth and sporulation
2.7 光照条件对寄生曲霉生长的影响
光照对寄生曲霉Q527生长的影响结果(图7)表明,菌丝生长受光照条件的影响差异显著。全黑暗条件下,菌丝生长速度最快,菌落直径为78.18 mm;光暗12 h交替下生长速率居中,菌落直径为76.04 mm;连续光照条件下生长速率最低,菌落直径为40.21 mm。产孢量受光照条件的影响差异显著,黑暗条件可促进寄生曲霉Q527产孢,产孢量为2.63×109个/皿。在不同光照条件下,持续光照对菌株产孢有抑制作用,但是产孢时间较早。
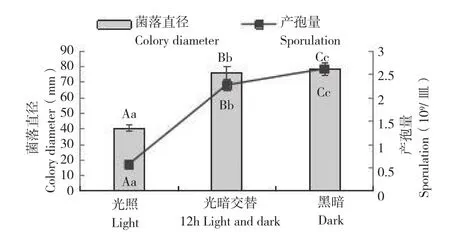
图7 不同光照条件对寄生曲霉生长和产孢量的影响Fig. 7 Effects of different light conditions on mycelial growth and sporulation
3 讨论
虫生真菌资源极为丰富,而我国已发现的昆虫病原真菌涉及40多个属400种[16]。白僵菌[17]、绿僵菌[18]和曲霉菌是自然界重要的虫生真菌资源,在生物防治中很早就被发现和利用。前人通过大量研究认为曲霉属的多种曲霉可作为生防菌可以用来防治目的昆虫,如采用烟曲霉和葡萄曲霉的融合菌株对松墨天牛越冬幼虫有较高的致死率[19]、米曲霉可抑制小菜蛾取食量减少并导致定殖率下降[20]、黄曲霉和溜曲霉对瓜实蝇成虫有较高毒性及防治效果[21]等。本研究首次报道寄生曲霉Q527对黄野螟幼虫具有良好的感染作用并在短期内导致黄野螟幼虫大量死亡。在病发虫体上发现黄霉层,可能是寄生曲霉直接侵染和次生代谢产物致病相互作用的结果[22]。曲霉生长发育除了对环境有更高要求外,营养元素的细微变化也会直接且迅速影响菌丝生长[23]。曲霉属不同种和不同菌株的最适外界环境和营养条件有所不同[24],本试验结果也说明了培养基、pH、碳源、氮源和光照的差异对曲霉Q527的生长和产孢量存在较显著影响。寄生曲霉在防治农林害虫具有十分明显的优势,但是生物防治的前提是针对不同菌株的特定培养条件进行培养,这需要对不同菌株的外界培养环境和内在营养条件进行研究,以期达到最大产孢量,这也是真菌大规模生产的要求[25]。随着新型技术的日渐成熟,有助于此菌种大规模在林间推广[26]。
4 结论
本研究结果表明,黄野螟幼虫的寄生曲霉是一种潜在的生防真菌,也是控制害虫的一种有效手段,其引起部分致病效应加快了昆虫的致死进程。寄生曲霉Q527生长最适培养基为PDA培养基,在pH5.0~12.0范围内均可生长。乳糖能促进寄生曲霉Q527菌丝生长,可溶性淀粉可有效促进寄生曲霉Q527产孢,蛋白胨作为氮源在生长速度和产孢量都有一定优势,营养繁殖喜黑暗,黑暗环境能促进其产孢。虽然研究结果表明此曲霉具备防除害虫的潜力,但对有前途的菌株等开展定性的安全性评价也很有必要,实际的开发利用还有待于进一步开展安全性研究。
