不同光质对竹柏幼苗叶片光合作用的影响*
2019-03-30游云飞
游云飞
(福建省洋口国有林场,福建顺昌353200)
光是植物生长发育的基本原因之一。光质对植物的生长、形态建成、光合作用、物质代谢以及基因表达均有调控[1]。不同植物种类、发育时期、生长状态、组织或器官对同一光质的反应不同,表现为光质生物学反应的复杂性[2]。研究表明,光质对植物光合作用具有重要的影响。红光可以通过抑制叶片中光合产物的输出来增加叶片中淀粉的积累,从而影响光合器官的发育,蓝光则调控叶绿素形成、叶绿体发育与气孔开启以及光合节律等生理过程[3];光敏色素、隐花色素、原叶绿素酸酯以及叶绿素均可调控叶绿体的发育,光敏色素主要感受红光与远红光,也感受蓝光与紫外光,隐花色素感受蓝光和UV-A[4];光质、光强还能够调节光合作用不同类型叶绿素蛋白质复合物的形成以及光系统II(PSⅡ)和光系统Ⅰ(PSⅠ)间的电子传递[5,6]。植物叶绿素荧光参数的变化可以在一定程度上反映环境因子的变化及其对植物光合生理产生的影响,其中,Fv/Fm用于表征PSⅡ反应中心光能的转化效率,非环境胁迫下该参数变化极小,为0.832±0.004,不受物种和生长条件的影响,但在逆境下这一比值明显降低[7]。因此,通过测定植物叶片叶绿素荧光参数Fv/Fm的变化,推测植物对特定环境因子的适应性和抗逆性可能是一条有效便捷的途径[8]。
竹柏(Podocarpusnagi),是罗汉松科(Podocarpaceae)竹柏属(Nageia)常绿乔木,是中生代白垩纪的孑遗植物[9,10],也是特殊的资源树种和优良的园林观赏树种[11]。对竹柏的研究目前主要集中在栽培[11]、种群结构[9]、形态结构[11]和次级代谢[12]等方面,尚未见对其光合生理的研究报道。发光二级管(light-emittingdiode,LED)作为新型半导体光源,具有光质纯、光效高、体积小、波长固定、发热低、冷却负荷小等优点,便于集中所需波长实施均衡近距离照射,是代替荧光灯用于植物设施栽培的新一代节能环保型光源[13]。本研究通过LED精确调制不同波长光质,探讨红光、白光和蓝光对竹柏幼苗光合作用和叶绿素荧光的影响,以期探讨出适合竹柏幼苗生长的光质条件,为竹柏栽培提供理论基础。
1 材料与方法
1.1 供试材料与试验设计
试验于福建农林大学田间实验室进行,供试材料来自武夷山金竹柏有限公司,选择生长一致、健壮、无病虫害的一年生轻基质竹柏容器苗。本试验选用红(波长662nm)、蓝(波长464nm)和白(波长454nm)三种光质,将上述植物材料置于LED培养箱内,该培养箱由台湾海博特股份有限公司生产,型号为HiPoint SC-3D、HiPoint SC-600。为了试验顺利进行,在预试验中选择红、蓝、白三种光质,30、60、90cd四种光强,每种组合40株幼苗,3个重复,观测光质、光强对苗木成活率及生长的影响;根据预试验,选用适宜植物生长的光强60cd,温度25±1℃,光周期8:00—20:00为12 h·d-1;每组12株,设3个重复,培养60 d;每组取3株竹柏幼苗,3个重复测定竹柏相关光合参数、叶绿素荧光参数和光合色素含量。

图1 LED光质生长培育光源图
1.2 测定项目与方法
在培养第60d测定光合参数、叶绿素荧光参数和光合色素含量。
光合参数采用LI-6400型便携式光合作用测定仪测定,测定时间为8:00—20:00,每隔2 h测定一次,每次测定3株植物,测定不同光质处理下植物净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)。
叶绿素荧光参数使用PAM2500便携式调制叶绿素荧光仪进行测定[14],实验开始前,将竹柏叶片放到暗夹中暗适应一段时间,测出初始荧光(Fo),然后给一个持续时间为0.5s的饱和脉冲,等到脉冲关闭时,测出竹柏的最大荧光(Fm),打开内源光化光,测出Fs,照射饱和脉冲光,测出Fm′;打开远红光,测出Fo′。根据以上数据,计算出PSII原初光能转化效率(Fv/Fm)、光下叶片实际光能转化效率(YII)、光化学淬灭系数(qp)、非光化学淬灭系数NPQ,每处理重复3次。
光合色素含量主要测定竹柏的叶绿素a、叶绿素b和类胡萝卜素含量,选取竹柏功能叶片进行测定,每个处理重复3次。采用“95%乙醇法”[15]测定光合色素含量。
1.3 实验数据与分析
实验数据采用Excel2010软件进行处理,用SPSS19.0进行方差分析。
2 结果与分析
2.1 对光合作用的影响
2.1.1 对净光合速率的影响
如图2所示,不同处理竹柏净光合速率日变化曲线差异较大,12:00前,白光和蓝光处理下的竹柏呼吸作用大于光合作用,净光合速率为负值,二者净光合速率日变化曲线相差不大,红光处理下的竹柏呼吸作用小于光合作用,净光合速率为正值。红光处理竹柏叶片净光合速率值与白光和蓝光处理变化相反,14:00时,白光、蓝光净光合速率达到第一个峰值,红光处理为谷值,蓝光比红光净光合速率高771.43%;白光和蓝光净光合速率于16:00达到谷值,均为 0.35 μmol·m-2·s-1,18:00 蓝光处理达到第二个峰值,为 1.10 μmol·m-2·s-1。白光和红光处理下光合速率日变化单峰曲线,蓝光处理为双峰曲线。
2.1.2 对蒸腾速率的影响
如图3所示,蒸腾速率变化与净光合速率变化趋势相反,白、红、蓝3种光质LED培养下的竹柏叶片蒸腾速率均为单峰曲线,红光处理的蒸腾速率变化趋势与白光和蓝光处理变化相反。14:00前,白光和蓝光蒸腾速率持续下降,红光处理蒸腾速率先下降后升高,并在10:00达到谷值,为0.15 mmol·mol-2·s-1,14:00达到峰值,为0.47 mmol·mol-2·s-1,白光和蓝光蒸腾速率在18:00达到峰值,但白光比蓝光蒸腾速率高22.86%,18:00后3个处理竹柏叶片蒸腾速率均下降。

图2不同光质处理下净光合速率日变化曲线

图3 不同光质处理下蒸腾速率日变化曲线
2.1.3 对气孔导度的影响
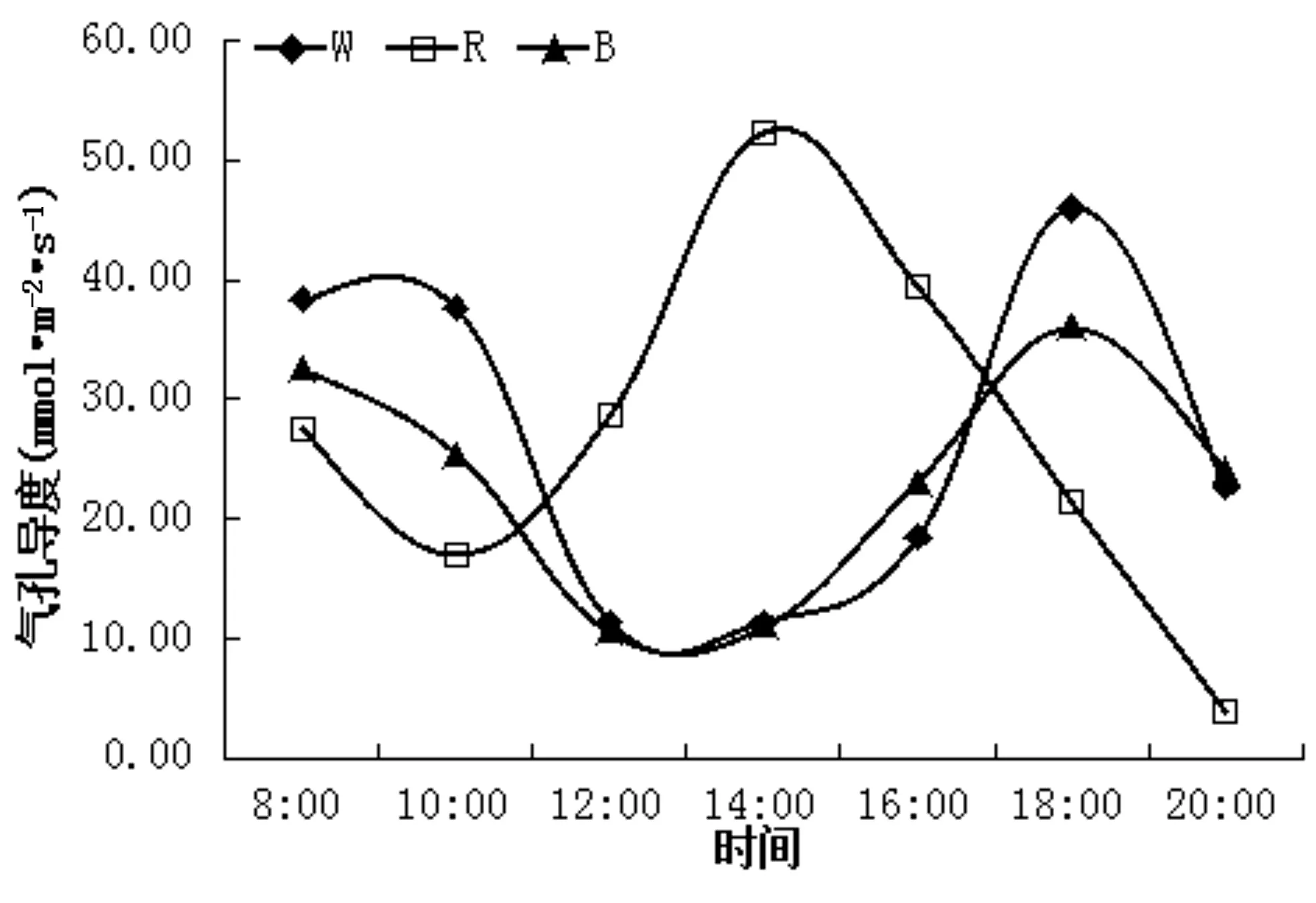
图4不同光质处理下气孔导度日变化曲线

图5 不同光质处理下胞间CO2浓度变化曲线
如图4所示,白光和蓝光气孔导度日变化趋势相似,红光与二者之间相差较大。10:00时红光处理竹柏叶片气孔导度达到谷值,为 17.00 mmol·mol-2·s-1,随后上升,14:00 时达到峰值,为 52.33 mmol·mol-2·s-1,比白光气孔导度高 361.87%,白光和蓝光处理在 18:00 气孔导度达到峰值,分别为 46.00 mmol·mol-2·s-1和 36.00 mmol·mol-2·s-1,18:00后3个处理竹柏叶片气孔导度均下降。
2.1.4 对胞间CO2浓度的影响
如图5所示,红光处理竹柏叶片胞间CO2浓度曲线变化与白光和蓝光处理相差较大,10:00红光处理竹柏叶片胞间CO2浓度在达到谷值,为225μmol·mol-1,比白光处理低53.22%,14:00白光和蓝光处理胞间CO2浓度达到谷值,分别为310.33μmol·mol-1和278.00μmol·mol-1,此时红光胞间CO2浓度达到峰值,比白光处理高23.85%,18:00后胞间CO2浓度均呈现下降趋势。
2.2 对光合色素含量的影响
如表1所示,不同光质培养影响竹柏叶片光合色素的含量。3个处理叶绿素a含量差异不显著,但白光处理竹柏叶片叶绿素a最大;白光和红光培养的竹柏叶绿素b含量显著高于蓝光处理,白光处理比蓝光高50.94%,白光和红光处理叶绿素b含量差异不显著;红光处理叶绿素含量介于白光和蓝光处理之间,白光处理叶绿素含量比蓝光处理高28.43%;叶绿素a/b值大小为:蓝光>白光>红光,3个处理相互间呈显著差异,蓝光处理下竹柏叶绿素a/b比红光处理高37.00%,比白光处理高26.85%。类胡萝卜素含量方面,3个处理含量没有显著差异,白光和蓝光培养下竹柏的类胡萝卜素含量相同,并且均大于红光处理。

表1 光质对竹柏光合色素含量的影响
2.3 对叶绿素荧光参数的影响
如表2所示,不同光质处理下竹柏叶片的叶绿素荧光参数产生变化。白光处理Fo和Fm最大,分别比蓝光大18.99%和15.49%,红光处理下竹柏叶片Fo与白光处理和蓝光处理没有显著差异,白光处理Fm显著高于红光处理和蓝光处理;3个处理Fv/Fm都为0.81,该条件下生长的竹柏均未受到光胁迫,各处理之间竹柏叶片的Y(II)、qp、NPQ差异不显著,但蓝光处理的NPQ较大,比红光处理大116.90%。

表2 光质对竹柏叶绿素荧光参数的影响
3 讨论
3.1 不同光质对竹柏幼苗叶片光合色素的影响
光合色素起到吸收、传递和转化光能的作用,是光合作用的基础[16],叶绿素和类胡萝卜素作为主要光合色素在植物光合作用中起到不同的作用,对光合作用影响较大。叶绿素可以有选择地吸收光能,是植物光合作用的光敏催化剂,叶绿素含量和叶绿素a与叶绿素b比例是衡量植物对环境适应性的重要指标。本试验中,叶绿素含量大小为:白光>红光>蓝光,与杜洪涛[17]研究的蓝光处理下彩色甜椒幼苗有较大叶绿素含量的结论不一致,可能与植物的种类和发育时期有关,表现出光质生物学反应的复杂性。叶绿素a和叶绿素b是叶绿素的重要组成部分,叶绿素a偏向吸收红光区的光,叶绿素b偏向吸收蓝光区的光[18]。不同光质培养下竹柏幼苗叶绿素a含量为:白光>蓝光>红光,叶绿素b含量为:白光>红光>蓝光,红光促进竹柏叶片中叶绿素b的形成,蓝光促进叶绿素a的形成,这与储钟稀、蒲高斌[19]等人研究结果一致。叶绿素a/b值可以作为植物对光环境适应的指标,叶片叶绿素a/b值约为3[20],不同类型植物叶绿素a/b值有所差异,阳生植物叶绿素a/b值较高,阴生植物叶绿素a/b值较低,3种光质处理下竹柏幼苗叶片叶绿素a/b值均低于3。类胡萝卜素起到吸收辅助色素、清除自由基、吸收和传递电子的作用,是光合作用中的辅助色素[21],红光处理类胡萝卜素含量最低,比白光和蓝光处理低20%,这与Anna和Alicja[22]等发现的红光降低风信子愈伤组织中类胡萝卜素含量的结果相同。白光和蓝光处理竹柏幼苗类胡萝卜素含量较高,光质对竹柏幼苗叶片类胡萝卜素含量的影响说明光敏色素控制了类胡萝卜素的合成,类胡萝卜素的增加可能对叶黄素的循环进行调控,保护光合器官免遭破坏[23]。
3.2 光质对竹柏幼苗叶片光合特性的影响
光合速率是影响植物同化能力和产量的关键因素,由于光合作用受到植物本身和外界条件影响较大,所以植物的光合速率一天中也存在着变化。蓝光培养的竹柏幼苗净光合速率日变化曲线为双峰曲线,白光和红光培养为单峰曲线,无光合“午休”现象。光合“午休”现象广泛存在于自然植物中,是植物普遍具有的现象。引起光合“午休”现象主要因素包括生理因子(生理节奏、气孔导度等)、生化因子(光化学系统、光合产物积累等)和生态因子(光照、温度、湿度等)[24]。相关研究表明,净光合作用日变化单峰曲线有利于提高竹柏的生物学产量[25]。竹柏幼苗蒸腾速率日变化与净光合速率日变化基本相同,蒸腾速率主要受到气孔的调控作用,光质与竹柏幼苗的蒸腾速率没有显著相关性,白光处理下竹柏幼苗的蒸腾速率比蓝光处理下的大。气孔控制外界CO2的进入和叶片内水分的散失,是植物与自然环境进行物质交换的通道,常用气孔导度表示气孔的开闭程度,气孔导度影响植物的光合作用和蒸腾作用[26-27]。胞间CO2浓度常用来判断光合速率变化的原因,即是否由气孔因素引起[28],胞间CO2浓度受到气孔导度、外界CO2浓度和叶片光合作用消耗的影响[29],不同LED光质处理中,红光处理胞间CO2浓度较低。根据Farquhar和Sharkey[30]提出的理论,判断光合速率降低由气孔因素引起的条件是,胞间CO2浓度和光合速率同时降低,且气孔限制值增大。由此可知,竹柏净光合速率的降低是由非气孔限制因素引起的,可能与叶肉细胞的光合活性有关。
3.3 光质对竹柏幼苗叶片PSⅡ活性的影响
植物能适应不同的光质和光强处理,得益于光合系统的多个调节水平和光合电子传递链各组份的协同作用。白、红、蓝3种光质处理中,白色LED培养的竹柏幼苗叶片Fo最高,比红光处理高11.90%,Fo可以反应PSⅡ反应中心受伤害情况,常用来判断PSⅡ反应中心的运转情况,这意味着与红光相比,白光培养的竹柏幼苗光合作用中PSⅡ反应中心遭到破坏,植物通过Fo的升高来缓解体内D1蛋白降解和活性氧的增加,进而保护PSⅡ反应中心,减少植物的伤害[31],此外,Fo代表光合色素吸收的光能中不参与光反应的部分,Fo升高说明用于光反应中的光能减少,多余的光能常以热和荧光形式散失[32]。其中白光培养的竹柏幼苗叶片叶绿素和类胡萝卜素含量较高,Fo也明显升高,说明PSⅡ反应中心受到破坏,导致白光处理下竹柏净光合速率较低。Fm表示PSⅡ反应中心电子传递情况,是植物暗适应一段时间后的最大荧光产量,PSⅡ反应中心完全关闭时,白光LED培养下竹柏的Fm最大。Fv/Fm是PSⅡ反应中心光能转换效率,反应植物受到光抑制的程度,正常情况下Fv/Fm值介于0.75到0.85之间,而且比较稳定。3种LED光质处理下竹柏幼苗叶片的Fv/Fm稳定,都为0.81,说明3种处理下的竹柏并没有产生光抑制现象。Y(Ⅱ)是植物叶片实际光能转化效率,表示线性电子在PSⅡ和PSⅠ间传递的量子效率,qp用来反映PSⅡ反应中心开放数目和原初电子受体qp氧化还原状态,qp越大,PSⅡ电子传递活性越大,NPQ主要是保护光合机构,反映PSⅡ天线色素吸收的能量中以热形式散失的光能。白光、红光和蓝光3种光质并没有对竹柏幼苗叶片光合反应中Y(Ⅱ)、qp、NPQ产生显著影响。
4 结论
(1)白光和红光培养的竹柏幼苗净光合速率日变化曲线为双峰曲线,无光合“午休”现象,说明红光和白光有利于竹柏提高竹柏生物产量。
(2)在红光处理下,竹柏幼苗叶片叶绿素a/b值和类胡萝卜素含量最低,而叶绿素a/b值在蓝光处理下最高。
(3)类胡萝卜素在蓝光和白光处理下最高,叶绿素含量和Fo值在白光处理下最高,说明白光处理下,竹柏幼苗PSII反应中心会遭受破坏。
