陕西杨凌地区番茄斑萎病毒外壳蛋白基因的克隆与分析
2019-03-14宁石岩秦蕾李云洲梁
默 宁石 岩秦 蕾李云洲梁 燕*
(1西北农林科技大学园艺学院,陕西杨凌712100;2贵州大学农学院,贵州贵阳550025)
番茄斑萎病毒(Tomato spotted wilt virus,TSWV)属于布尼亚病毒目(Bunyaviridae)番茄斑萎病毒科(Tospovirus)正番茄斑萎病毒属(Orthotospovirus),被列为世界十大植物病毒之一(Rosel ló et al.,1996;Ullman et al.,2005),最早于1915年在澳大利亚被发现(Brittlebank,1919)。该病毒寄主植物广泛,可侵染84个科1 090种植物。TSWV可系统侵染整株植株,进而导致发病。该病毒的传播介体主要是西花蓟马以持久循环增殖型传播方式传播,传播介体一旦感染该病毒,便会终身带毒(郑雪 等,2013;李云洲 等,2018)。TSWV已在巴西、阿根廷、法国、西班牙、意大利、澳大利亚、美国等国家的番茄生产中造成了严重损失(Jones,2005)。我国于2003年检测出TSWV(Soler et al.,2003),之后随着TSWV危害范围逐年扩大,在四川、云南、广州、山东、河北等地多种园艺植物上均有发现(丁铭 等,2004;张友军 等,2004;Dong et al.,2008;饶雪琴 等,2010;方琦 等,2011;邱树亮 等,2012;郑雪 等,2013)。目前,TSWV在云南等地分布广泛,危害也日趋严重,大棚和温室番茄均有受损,严重时产量损失超过80%,甚至绝收(方琦 等,2011)。
陕西杨凌是我国唯一一处国家级农业高新技术产业示范区,番茄为杨凌地区的主要蔬菜作物。为了了解TSWV在杨凌地区的发病情况以及致病病毒类型,于2016年4~7月在番茄主要栽培示范基地进行调查取样,收集了疑似感染TSWV的番茄样品4份,进行分子鉴定,明确杨凌地区TSWV病毒株系,为杨凌地区番茄TSWV鉴定以及抗TSWV育种提供依据。
1 材料与方法
1.1 试验材料
1.1.1 植物材料 选取杨凌地区感病番茄植株上部幼嫩叶片,锡箔纸包裹后置液氮冷冻5 min后于-80℃超低温冰箱中保存备用。
1.1.2 载体与菌株 pMD18-T购于大连宝生物工程有限公司。大肠杆菌(Escherichia coli)感受态DH5α购自天根生化科技(北京)有限公司。
1.2 试验方法
2016年4~7月于杨凌区新天地设施大棚中设点观察番茄感染TSWV的症状。采集疑似感病材料发病叶片,液氮速冻,提取叶片RNA,利用TSWV外壳蛋白基因(CP)的特异性引物(F:5′-TCTGTGAGGCTTGCCATAAT-3′;R:5′-A GCATACTCTTTCCCTTTCT-3′)对田间疑似发病叶片进行PCR鉴定。
1.3 杨凌地区TSWV CP基因克隆测序及分析
利用已检测出的阳性材料进行TSWV CP基因的全长克隆,PCR扩增引物为F:5′-ATGTYTAAGGTTAAGCTCACTAAG-3′;R:5′-TT AAGCAAGYYCTGYGAGTTTTGCC-3′,将 PCR 产物送往西安擎科生物公司进行测序,利用DNAMAN 6.0软件对测序所得序列进行比较分析;为了分析杨凌地区TSWV番茄分离物系统进化关系及分类地位,与已有的TSWV分离物,基于TSWV CP基因序列,采用MEGA 5.0软件以邻接法构建系统发育进化树,各分支置信度(bootstrap)进行1 000次重复分析。
2 结果与分析
2.1 番茄TSWV的田间症状观察
番茄感染TSWV后,初期叶片开始有不太明显的深色病斑(图1-B),随着病斑面积的扩大,叶片开始发黄坏死;进而植株的茎上也出现病斑(图1-A),开始时期零星分布,随着症状的发展,病斑连成一片,整棵植株长势越来越弱;绿熟期果实出现明显病斑,表面凹凸不平(图1-C),红熟期果实也有古铜色斑点出现(图1-D)。

图1 杨凌地区设施番茄感染TSWV后的症状表现
2.2 杨凌地区番茄TSWV鉴定结果
用TSWV CP基因的特异性引物对4份供试样品进行PCR检测,其中2份样品扩增到约234 bp的特异性条带(图2-A),经测序后比对,这2份阳性样品扩增出的特异条带序列与GenBank中的TSWV序列相似性均达到98%以上,表明所扩增到的基因片段属于TSWV。
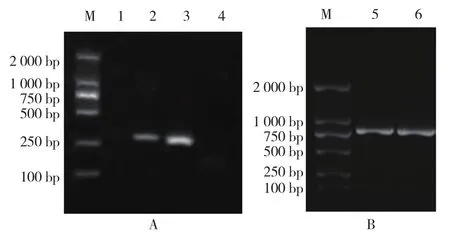
图2 杨凌地区TSWV鉴定(A)及其CP基因的克隆(B)
2.3 杨凌地区TSWV CP基因的克隆与测序
选取TSWV检测为阳性的番茄样品,采用同源克隆方法扩增TSWV的CP基因全长,得到目的片段(图2-B),并对目的片段进行回收,连接pMD-18T,热激转化进大肠杆菌(Escherichia coli,Top10),测序后获得757 bp的TSWV CP基因(图3),ATG为起始密码子,TAA为终止密码子。
2.4 杨凌地区TSWV CP基因序列的同源性分析

图3 杨凌地区TSWV CP全基因序列
从表1可以看出,杨凌地区TSWV与西班牙TSWV(FR693089.1)CP基因的同源性最高,达到99%;与阿尔及利亚TSWV(FR693045.1)的同源性为90%;与夏威夷TSWV(X61799.1)、中国台湾(HM180089.1)、美国(HQ406983.1)、澳大利亚(AY879109.1)、新西兰(KC494503.1)、土耳其(KM379142.1)、中国云南(KC294570.1)的同源性在79%~86%之间;而与中国昆明TSWV(HQ402595.1)、 韩 国(HM581936.1)、 朝 鲜(KC261949.1)的同源性仅为15%~16%。
从氨基酸序列比对结果可以看出(图4),除 了 中 国 昆 明 TSWV(HQ402595.1)、 韩 国(HM581936.1)、朝鲜(KC261949.1)外,杨凌地区TSWV与其余9个地区的TSWV氨基酸序列的保守区域基本一致。跨膜分析结构显示(图5),杨凌地区TSWV的外壳蛋白具有1个跨膜结构,位于95~122个氨基酸之间。
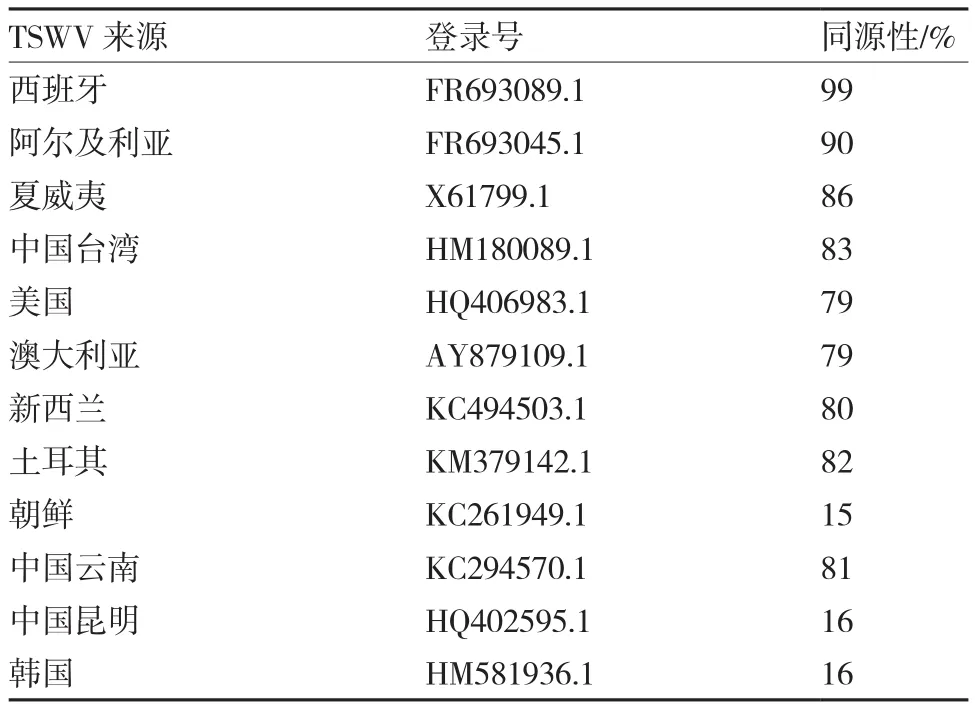
表1 杨凌地区TSWV与NCBI中12个TSWV CP基因序列的同源性

图4 13个TSWV外壳蛋白氨基酸序列比对结果
2.5 系统进化树分析
系统发育进化树显示(图6),杨凌地区番茄样品的TSWV序列与西班牙TSWV分离物(FR693089.1)处于同一分支,亲缘关系最近,并且二者与阿尔及利亚分离物(FR693045.1)处于同一大支,亲缘关系较近;而杨凌地区TSWV番茄分离物与云南昆明的TSWV分离物亲缘关系较远,昆明TSWV分离物与韩国TSWV分离物的亲缘关系较近。

图5 TSWV蛋白跨膜区的预测结果
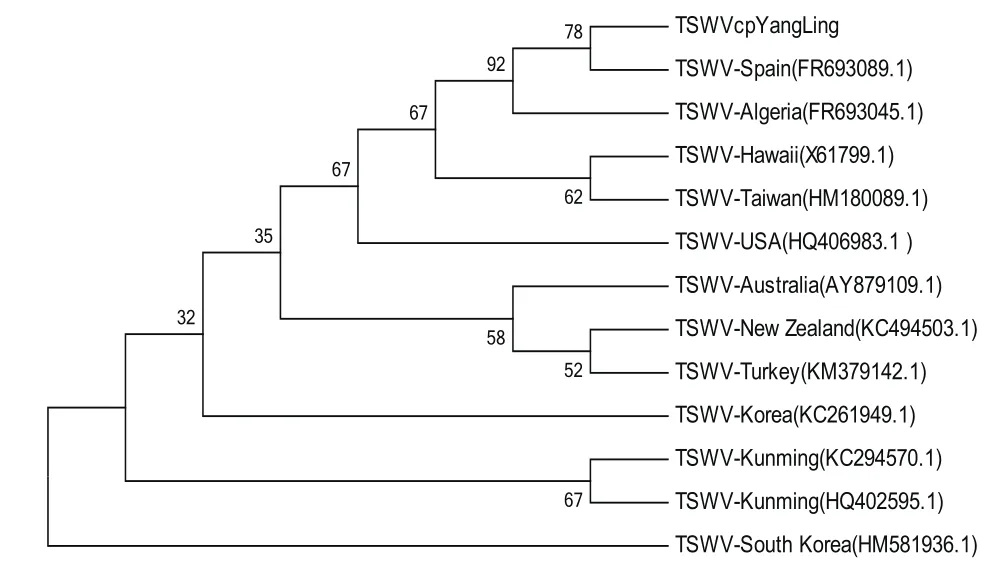
图6 基于CP基因序列构建的杨凌地区TSWV分离物及其相关分离物的系统进化树
3 结论与讨论
TSWV在我国出现较晚,但随着西花蓟马的传播,危害范围逐渐扩大,危害也日趋严重(方琦 等,2011),由于地理环境差异,该病毒种类多,亲缘关系复杂。目前关于番茄抗TSWV的标记有9个,分别为 Sw-1a、Sw-1b、Sw-2、Sw-3、Sw-4、Sw-5、Sw-5b、Sw-6、Sw-7(Finlay,1952,1953;van Zijl et al.,1986;Stevens et al.,1991;Canady et al.,2001;Lee et al.,2015),在育种中应用较多的为Sw-5。由于病毒NSs基因突变可以造成Sw-5抗性消失(Jiang et al.,2017),因此了解当地病毒基因序列十分关键。本试验通过分子鉴定获得了杨凌地区TSWV的CP基因序列,为当地番茄抗病育种提供了基础。
国内外已有不少关于番茄斑萎病毒基因克隆及序列比对分析的研究(方琦 等,2011;尹跃艳 等,2013),尹跃艳等(2013)对21个TSWV昆明分离N基因序列进行克隆分析,系统进化分析发现昆明分离物与韩国分离物的亲缘关系很近,而本试验中杨凌地区的TSWV分离物CP基因与西班牙番茄上的TSWV分离物亲缘关系更近,而与昆明的TSWV分离物的亲缘关系较远。可见不同地区的TSWV存在差异。
当前TSWV在我国各地普遍发生,而我国对于TSWV的研究仍处于起步阶段。应首先从研究当地TSWV病毒类型入手,选育适应当地TSWV病毒类型的抗病品种,挖掘新的抗病基因,增加番茄抗TSWV基因的储存量,使抗病基因多样化以适应不同地区的TSWV病毒类型,这对我国番茄TSWV抗性育种工作的开展具有积极意义。
