汉丰湖湖岸水敏性系统鸟类多样性
2019-02-26袁兴中刁元彬张乔勇
刘 恋 ,袁兴中 *,刘 红 ,刁元彬 ,张乔勇
(1.煤矿灾害动力学与控制国家重点实验室,重庆 400030;2.重庆大学 资源及环境科学学院,重庆 400030)
近年来,水敏性城市设计(water sensitive urban design)作为具有综合功能的全新水管理方式,逐渐成为城市化研究的热点。相关研究侧重于水敏性系统削峰滞流、污染净化、水资源利用等功能[1-3]。随着城市化进程的加速,原有生境往往被人工生境不可逆替代,导致生境数量及质量急剧下降。城市绿地在面积大小、生境异质性、生境连续性、植被结构上均劣于原有生境,导致城市化过程中生物多样性降低[4]。水敏性系统作为城市绿地的全新模式,除了水环境管理功能外,在维持、提升城市生物多样性上具有重要作用[5],但相关研究较少。
鸟类作为城市生物多样性的重要组成部分,具有对环境变化敏感的特点,是识别、了解城市生物多样性现状,评价城市生态系统健康水平的重要生物指标[6-7]。国外有关城市鸟类生态学的研究较多[8-9],近年来国内陆续开展了城市化进程及不同类型城市绿地对鸟类多样性影响的研究[10-12],但针对城市鸟类多样性提升的城市绿地设计及研究较少[13-14],水敏性系统维持鸟类多样性的研究仍为空白。
笔者以处于快速城市化区域的汉丰湖湖岸水敏性系统为例,开展了以鸟类作为生物指标的城市生物多样性调查,以期定量评估水敏性系统维持、提升城市生物多样性的功能,探讨水敏性系统维持城市生物多样性的机制,为城市生物多样性保育及水敏性系统结构、功能优化及推广提供科学依据。
1 研究区域与研究方法
1.1 研究区域概况
汉丰湖位于三峡库区腹心的重庆市开州区新城北部,属长江一级支流澎溪河的回水末端。汉丰湖区域属亚热带季风气候,多年平均气温18.5℃,多年平均降水量1 385 mm。
汉丰湖湖岸共设置两个样点(图1),样点一为水敏性系统(以下称为实验区),是室内外一体的整体生态系统设计,面积为2 480 m2,水敏性系统集“生命景观屋顶-生命景观墙-雨水花园”为一体,具有消纳城市地表径流、净化水质、提升城市生物多样性等功能。其中生命景观屋顶面积为183.4 m2,主要种植的植物为肾蕨(Nephrolepis auriculata)、天门冬(Asparagus cochinchinensis)、马缨丹(Lantana camara)、墨西哥鼠尾草(Salvia leucantba)、黄槐决明(Cassia surattensis)、芒(Miscanthus sinensis)等。生命景观墙总面积为252.9 m2,主要种植的植物为天门冬、肾蕨、鹅掌柴(Schefflera octophylla)、沿阶草(Ophiopogon bodinieri)等。雨水花园总面积为2 018.3 m2,主要种植植物为樟(Cinnamomum camphora)、皂荚(Gleditsia sinensis)、黄葛树(Ficus virens)、萼距花(Cuphea hookeriana)、女贞(Ligustrum lucidum)、马樱丹、沿阶草、天门冬、肾蕨等。雨水花园包括生物沟、下沉式绿地、浅凹绿地等下渗系统,以及蜻蜓塘、青蛙塘等小微湿地,小微湿地约为2 m×4 m,形状以椭圆形、弯月形为主,蓄水深度为20~40 cm,以水生植物为主,包括石菖蒲(Acorus tatarinowii)、泽泻(Alisma plantago-aquatica)、鸢 尾(Iris tectorum)、黄花水龙(Ludwigia peploides)等小型挺水植物,荇菜(Nymphoides peltatum)、萍蓬草(Nuphar pumilum)等浮叶根生植物。样点二(以下称为对照区)为实验区西侧270 m的人工草坪,面积为2 540 m2,乔木优势种为日本晚樱(Cerasus serrulata)、木犀(Osmanthus fragrans),灌木优势种为红花檵木(Loropetalum chinense)、山茶(Camellia japonica)、女贞,草本植物优势种为狗牙根(Cynodon dactylon)、细叶结缕草(Zoysia tenuifolia)、马兰、白车轴草(Trifolium repens)、莲子草(Alternanthera sessilis)。二者同为城市绿地,具有面积小、斑块化、人为干扰强烈等特点。

图1 实验区(左)与对照区(右)概貌Fig.1 The general appearance of experimental area(left)and control area(right)
1.2 研究方法
1.2.1 采样方法
2017年5月、8月、10月分别对实验区和对照区进行鸟类调查,采用样点法,每次调查3 d。选择晴朗无风的天气,在鸟类活动的高峰段(6:30-8:30和16:30-18:30)进行调查,每天重复两次。每次调查,在实验区和对照区分别观察20 min。调查人员携带8×42倍双筒望远镜及长焦相机进行观察和拍照,记录鸟类种类、数量、行为、生境、人为干扰状况等信息。鸟类辨识参照《中国鸟类野外手册》[15],分类参照《中国鸟类分类与分布名录》[16]。
1.2.2 数据分析
根据鸟类群落的特点及采样数据,选择以下模型计算[17-18]。

式中:Ni为第i个物种的个体数量;N为总个体数;S为物种数;Pi为第i种个体数量占群落总个体数量的比例;j为实验区与对照区份共有的鸟类种类数;a为实验区的鸟类种类数量;b为对照区的鸟类种类数量。其中优势度等级按照如下标准划分:优势种(J>10%),常见种(1%≤J≤10%),稀有种(J<1%)[19]。相似程度按照如下标准划分:极相似(0.75≤c<1),中等相似(0.25≤c<0.75),中等不相似(0.15≤c<0.25),极不相似(c<0.15)。
用One-way ANOVA对鸟类群落数量指标进行差异显著性检验,若差异显著,采用Duncan法进行多重比较,数据采用SPSS 20.0统计软件进行统计处理。
1.2.3 功能群划分
参考尚玉昌[20]、赵正阶[21]、Paszkowski等[22]的研究,并结合本研究鸟类取食习性的观察,将鸟类划分为4种取食功能群:(1)肉食性鸟,取食其他鸟类、小型哺乳动物等;(2)食虫鸟,主要取食昆虫、陆生节肢动物和某些环节动物;(3)植食性鸟,主要取食植物叶、嫩枝、果实、种子、根茎、块茎等;(4)杂食性鸟,取食小型脊椎动物、昆虫、植物体等多种食物。
2 结果分析
2.1 群落组成
2.1.1 种类组成
本研究共记录鸟类314只,隶属4目19科27种(表1),占汉丰湖区域鸟类97种的27.84%[23]。其中实验区共记录鸟类185只,隶属3目17科23种,鸟类种类数由多到少依次为雀形目(21种)、鴷形目(1种)、鸽形目(1种),雀形目种类最多,占总种类的91.30%。从数量上看,优势种为北红尾鸲、灰椋鸟、棕背伯劳,常见种为红头长尾山雀、白颊噪鹛、大山雀、黄腰柳莺等,少见种为红尾水鸲、树鹨、大拟啄木鸟、珠颈斑鸠等。对照区共记录鸟类129只,隶属2目11科15种,鸟类种类数由多到少依次为雀形目(14种)、鹤形目(1种),雀形目种类最多,占总种类的93.33%。从数量上看,优势种为纯色山鹪莺、白鹡鸰、白腰文鸟、北红尾鸲,常见种为大山雀、乌鸫、白头鹎、鹊鸲等,少见种为黑卷尾、红胸田鸡。
实验区留鸟最多,有19种,占总种数的82.61%;其次为夏候鸟2种、冬候鸟1种,分别占8.70%、4.35%;旅鸟较少,仅1种。对照区留鸟最多,有12种,占总种数的80.00%;其次为夏候鸟3种,占20.00%;无冬候和旅鸟(图2)。
2.1.2 区系分析
根据动物地理区系划分,将本研究观察到的鸟类划分为古北界、东洋界和广布种。实验区共记录到东洋界鸟类12种,占鸟类总种数的52.17%;广布种鸟类7种、古北界鸟类4种,分别占30.43%和17.39%。对照区共记录到东洋界鸟类7种,占鸟类总种数的46.67%;广布种鸟类6种、古北界鸟类2种,分别占40.00%和13.33%。研究区域为东洋界,实验区、对照区鸟类主要为东洋界,鸟类区系组成与研究区域所处区系一致。

图2 实验区与对照区鸟类留居类型对比Fig.2 The comparison of avian seasonal variation in experimental area and control area
2.1.3 生态类群
根据鸟类生态类群划分,将本研究观察到的鸟类划分为鸣禽、陆禽、攀禽和涉禽。实验区鸣禽占绝对优势,有21种,占鸟类总种数的91.30%;陆禽、攀禽均为1种。对照区鸣禽占绝对优势,有14种,占鸟类总种数的93.33%;涉禽仅1种。
2.2 鸟类群落季节变化
2.2.1 优势种
研究区域鸟类优势种种类随季节变化明显。实验区春季优势种为棕背伯劳、黑尾蜡嘴雀、丝光椋鸟,夏季为白颊噪鹛、大山雀、棕背伯劳,秋季为红头长尾山雀、灰椋鸟、北红尾鸲、白头鹎;对照区春季优势种为白鹡鸰,夏季为大山雀、乌鸫、纯色山鹪莺、白鹡鸰,秋季为白腰文鸟、北红尾鸲、纯色山鹪莺、白鹡鸰。
2.2.2 多样性的季节变化
实验区及对照区鸟类物种丰富度、种类及个体数量随季节变化明显(图3)。实验区秋季鸟类物种丰富度和个体数量均高于春季和夏季;春季与夏季物种丰富度相同,夏季个体数量略高于春季。实验区秋季鸟类物种丰富度及个体数量与春季、夏季差异显著(P<0.05);春季与夏季之间差异不显著(P>0.05)。对照区秋季鸟类物种丰富度及个体数量与春季差异显著(P<0.05),与夏季差异不显著(P>0.05);夏季与春季鸟类物种丰富度差异显著(P<0.05),个体数量差异不显著(P>0.05)。实验区与对照区之间鸟类物种丰富度春季、秋季差异显著(P<0.05),夏季不显著(P>0.05);秋季鸟类个体数量差异显著(P<0.05),春季、夏季不显著(P>0.05)。

表1 实验区及对照区鸟类种类组成与数量Tab.1 Species composition and quantity of avian community in experimental area and control area
实验区及对照区鸟类物种各多样性指数随季节变化明显。实验区鸟类Shannon-Wiener指数秋季最高,春季、夏季相等,各季节间差异不显著(P>0.05)。对照区鸟类Shannon-Wiener指数秋季与夏季显著高于春季(P<0.05),秋季与夏季差异不显著(P>0.05)。实验区各季节鸟类Shannon-Wiener指数均高于对照区,春季、秋季差异显著(P<0.05),夏季不显著(P>0.05)。实验区鸟类Pielou均匀度指数春季、夏季均显著高于秋季(P<0.05),春季与夏季差异不显著(P>0.05)。对照区鸟类Pielou均匀度指数规律与实验区相同。实验区与对照区之间各季节鸟类Pielou均匀度指数差异不显著(P>0.05)。

图3 不同季节实验区与对照区鸟类群落各多样性指数(均值±标准差)Fig.3 Species diversity index in different seasons of experimental area and control area(Mean±SD)
2.2.3 鸟类群落结构
实验区秋季与春季、夏季鸟类群落结构相似性系数低,鸟类群落结构组成差异性明显;春季与夏季鸟类群落结构组成中等相近,相似性系数约为0.5。对照区规律与实验区相似。对照区与实验区春季鸟类群落结构相似性系数低,鸟类群落结构组成差异性明显;夏季、秋季组成中等相似,相似性系数约为0.5。
2.2.4 鸟类取食功能群季节变化
实验区鸟类取食功能群相对丰富度和相对多度随季节变化规律基本一致(图4、5)。春季食虫鸟、杂食性鸟、植食性鸟和肉食性鸟相对丰富度和相对多度较为均匀;夏季、秋季以食虫鸟为主,杂食性鸟次之,夏季无植食性鸟。食虫鸟相对丰富度、相对多度从春季到秋季呈上升趋势,相对多度在秋季高达83.19%。肉食性鸟相对丰富度、相对多度从春季到秋季呈下降趋势,相对多度在秋季降低至2.65%。植食性鸟仅在春季、秋季出现,且秋季相对丰富度、相对多度低于春季。对照区鸟类取食功能群季节变化趋势与实验区相似,但秋季出现大量白腰文鸟等群居植食性鸟,极大提升了植食性鸟类相对丰富度和相对多度。
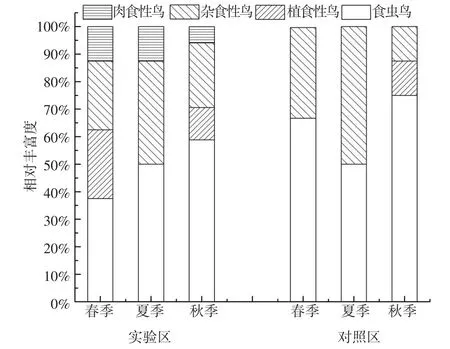
图4 实验区与对照区鸟类取食功能群相对丰富度的季节变化Fig.4 Temporal changes of avian feeding functional groups'relative richness in the different seasons of experimental area and control area

图5 实验区和对照区鸟类取食功能群相对多度的季节变化Fig.5 Temporal changes of avian feeding functional groups'relative abundance in the different seasons of experimental and control area
3 讨论
与自然环境相比,城市生态系统具有异质性高、污染严重、人类干扰频繁、生境破碎化等特点。多数学者认为,城市化进程导致城市生境破碎化、植被结构单一、小面积绿地斑块比重增加、连通性降低[24],从而降低鸟类多样性。而另一部分学者则认为,在适度的城市化压力下,城市中丰富的食物来源、较低的天敌捕食压力、城市中度干扰等因素使得鸟类多样性得以提升[25]。城市绿地作为城市生态系统的重要组成部分,为城市生态系统提供了诸多的服务功能,在维持、提高生物多样性方面有巨大潜力[26-30],是城市生态系统中鸟类重要的栖息地和避难所。
3.1 城市绿地生境异质性对鸟类多样性的保育功能
PALMER等[31]的研究表明,通过保持适当的栖息地和植被结构复杂性,能够有效维持城市鸟类多样性。对于陆生鸟类而言,乔木、灌木是其最为重要的生境因子,复杂、稳定的木本植物群落是其栖息、觅食、繁殖、躲避天敌的最佳场所[32-33]。相较于对照区而言,实验区木本植物丰富度、多度、盖度等更高,其复杂的植被结构和生境类型能够吸引更多的鸟类取食功能群,同时维持更高的多样性。除了城市绿地植被结构与组成对鸟类分布影响较大之外,生境异质性也是影响鸟类群落组成的重要因素,小微湿地、郁闭灌丛等形成的特殊微生境能够为一些对栖息地有特殊要求的鸟类提供适宜的生境[34]。实验区是集“生命景观屋顶-生命景观墙-雨水花园”为一体的立体绿地,与对照区相比,生命景观屋顶和生命景观墙中生长繁茂的草本和灌木成为了鸟类的庇护场所。此外,雨水花园内零星分布的塘改变了实验区整体环境的水分条件,影响其中昆虫的种类与数量,从而影响鸟类的食物结构,丰富食物来源。就城市公园等大尺度城市绿地而言,湖泊、水道、山丘等异质性生境有利于维持鸟类多样性[35]。林石狮等[36]的研究表明,快速城市化过程中保留的大型水库及其周围的绿地能够为游禽提供适宜的栖息地,可有效增加城市生物多样性。可见,植被组成越复杂、空间异质性越高的城市绿地能够提供更多的生态位,所支持的鸟类生物多样性越高。
3.2 食源对鸟类多样性的保育功能
对于鸟类而言,食物来源是影响其群落结构与分布的重要因素[37-38]。在城市绿地中,皱叶冬青(Ilex perryana)、火棘(Pyracantha fortuneana)、南天竹(Nandina domestica)、樟等常见的浆果植物能够为植食性、杂食性鸟类提供丰富的食源,适当提高观果树种的种类和比例,有利于城市中鸟类多样性的保护[14]。此外,对蝴蝶而言,实验区的马缨丹、萼距花、墨西哥鼠尾草、女贞等植物是典型的蜜源植物,苎麻(Boehmeria nivea)、黄槐决明(Cassia surattensis)、凹叶景天(Sedum emarginatum)、樟、小苜蓿(Medicago minima)、合欢(Albizia julibrissin)、堇菜(Viola verecunda)等是典型的宿主植物,有利于维持蝴蝶多样性,不同时期的鳞翅目幼虫、成虫是食虫鸟和杂食性鸟类重要的食物来源,可在不同季节满足金腰燕、黑喉石鵖、白头鹎、白颊噪鹛、棕背伯劳等鸟类的捕食需求,这也是实验区各季节鸟类物种丰富度、个体数量、Shannon-Wiener指数均大于对照区的原因之一。肉食性鸟虽仅棕背伯劳一种,但在实验区各季节均有出现,且伴随衔枝筑巢、捕食青凤蝶等行为。说明较对照区而言,实验区具有更稳定、丰富的食物来源,这是棕背伯劳等肉食性鸟类长期定居的必要条件。
3.3 鸟类多样性与城市生态系统健康
鸟类取食植物果实后,难以消化的种子随其粪便进入环境,间接完成了植物种子的传播。尤其在生境破碎化的城市区域,原本依靠小型哺乳动物进行的种子传播由于生境隔离、通道受阻而被极大削弱,而鸟类的活动空间相对自由,在城市生态系统中发挥着重要的种子传播功能,有利于植物群落的更新和新物种的引入,可促进植物多样性的提升。此外,食虫鸟和杂食性鸟能够消灭大量树木害虫,对于城市绿地的病虫害防治意义重大,是维持、促进城市生态系统健康发展的重要生物。
3.4 水敏性系统与城市生物多样性
水敏性系统作为城市生境的重要组成部分,与草地、花圃、人工次生林等传统城市绿地相比,通过立体绿化增加了单位面积的绿化率,具有植物丰富度,植被结构优于草地、花圃,生境更为复杂、异质,且用地面积远小于城市公园等人工次生林的优点。Kazemi等[5,29,39]在墨尔本的研究也证明,生物滞留池等水敏性系统中地表无脊椎动物生物物种丰富度、多度、多样性均显著高于普通花园、草坪等传统城市绿地,水敏性系统是提高城市生物多样性的有效途径。
在现有城市绿地规划及建设中,城市绿地往往植物种类及结构单一、生境异质性低,很少考虑到其地表径流污染净化、城市雨洪管理及生物多样性提升的功能,汉丰湖湖岸水敏性系统的建设正是针对以上问题开展的。开州区基于汉丰湖水质安全,开展了一系列城市生态绿地空间的探索,在汉丰湖湖岸进行了室内外一体的整体生态系统设计。本研究表明,与传统草坪绿地相比,水敏性系统能够有效维持、提高城市鸟类多样性,在城市规划中,应尽可能减少草坪等不可持续的城市绿地,扩大水敏性系统的应用。即使目前能证明水敏性系统实际生物多样性功能的研究较少,但水敏性系统仍是当前城市规划中最具生物多样性潜力的城市绿地模式[40-41]。
4 结论
(1)2017年5—10月,在汉丰湖湖岸选取水敏性系统(实验区)及对照区进行鸟类调查。共调查鸟类314只,隶属4目19科27种。实验区及对照区鸟类优势种种类随季节变化明显。
(2)各季节实验区物种多度、丰富度、Shannon-Wiener指数均高于对照区,且均在食源丰富的秋季达到最高值。实验区及对照区鸟类Pielou均匀度指数春季、夏季均显著高于秋季(P<0.05),但实验区与对照区之间各季节鸟类Pielou均匀度指数差异不显著(P>0.05)。
(3)实验区及对照区秋季与春季、夏季鸟类群落结构组成差异性明显,春季与夏季鸟类群落结构组成较为相似。
(4)实验区及对照区各季节鸟类摄食功能区均以食虫鸟为主,且食虫鸟相对丰富度及相对多度从春季到秋季呈上升趋势。
