基于Cyt b基因全序列鼩鼱属分子系统学分析—我国东北地区鼩鼱属物种的系统发生地位
2019-02-26张隽晟杜建宇金志民
张隽晟 刘 铸* 董 明 杜建宇 杨 茜 金志民,2
(1.牡丹江师范学院生命科学与技术学院,牡丹江,157011;2.东北林业大学野生动物资源学院,哈尔滨,150040)
鼩形目(Soricomorpha)鼩鼱科(Soricidae)具有2个亚科,即鼩鼱亚科(Soricinae)和麝鼩亚科(Crocidurinae),鼩鼱属(Sorex)隶属于鼩鼱亚科。鼩鼱属是鼩鼱亚科中种类最多的一个属,约70个物种,分布范围广泛,是鼩鼱科中唯一的既分布于古北区又分布于新北区的属[1]。Hoffmann认为我国具有该属14个物种[2],且在我国东北还可能存在2个物种,即扁颅鼩鼱(Sorexroboratus)和苔原鼩鼱(S.tundrensis),近些年,这两个物种在中国的分布已经得到证实[3-4]。Hoffmann在对我国鼩鼱亚科的系统分类和分布进行评论时,将我国鼩鼱亚科动物的分布划分为4个生物地理类群:东北寒温带针叶林(泰加林)、东北寒温带针叶林山区针叶林中具有孤立居群、中国喜马拉雅地区、中国特有种[2]。其中前两个生物地理类群具有的物种中,除了水鼩鼱(Neomysfodiens)外,其余分布物种都是鼩鼱属物种,因此我国东北分布的鼩鼱属种类属于前2个生物地理类群,是我国鼩鼱属的重要组成部分。刘铸等对我国东北地区进行鼩鼱科动物进行调查时发现,东北地区具有鼩鼱属9个物种:大鼩鼱(S.mirabilis)、中鼩鼱(S.caecutiens)、远东鼩鼱(S.isodon)、苔原鼩鼱、长爪鼩鼱(S.unguiculatus)、栗齿鼩鼱(S.daphaenodon)、细鼩鼱(S.gracillimus)、扁颅鼩鼱和姬鼩鼱(S.minutissimus)[5]。
鼩鼱属最早的化石纪录在中新世末和上新世初之间,由于该属的化石记录很稀少,并且确定化石的分类地位存在较大困难[6],因此,起初的鼩鼱属系统分化研究主要依靠形态学、核型和生化分析,这些方法都存在着各自研究的局限性[2,7],因此鼩鼱属的系统分化一直受到关注,诸多问题存在争议[8]。 随着分子生物学的快速发展,利用线粒体DNA(Mitochondrial DNA,mtDNA)序列对鼩鼱属的分子系统学研究,已经解决了一些鼩鼱属的系统分化和分类学问题。利用mtDNA序列的分子系统学研究主要集中在欧洲的西南部地区[9]、新大陆[10]和XY1Y2性染色体复杂种组(Sorexaraneus-arcticusgroup)[11]。近些年,东北亚地区的鼩鼱属系统分化问题得到了一定重视,但研究利用的mtDNA的序列长度有限,很多系统分化问题没能得到较好的解决[12-13]。Fumagalli等利用mtDNA的细胞色素b(Cytochrome,Cytb)基因较长的片段,对鼩鼱属的分子系统分化问题做了较为系统的研究,但研究建立在一些受争议的基础上,例如:将苔原鼩鼱归属于新北区物种,实际该物种为全北区物种,再有该研究涉及的物种有限,只对29个物种进行了分析,没有包括我国东北地区分布的大鼩鼱、栗齿鼩鼱、细鼩鼱和扁颅鼩鼱[8]。
本文期望利用mtDNA的Cytb基因全序列对39个鼩鼱属物种的系统分化进行分析,进而探讨我国东北地区具有的9个鼩鼱属物种的系统分化和分类学问题,为今后东北地区小型哺乳动物资源调查、群落结构分析和动物区系分化等生态学研究奠定一定基础。
1 材料和方法
1.1 标本采集
主要利用陷阱法,配有夹捕法,在大兴安岭山脉呼中地区(DH)、塔河地区(DT)和额尔古纳地区(DE);小兴安岭山脉萝北地区(XL);长白山山脉横道河子地区(CH)、三道关地区(CS)、牡丹峰地区(CM)、凤凰山地区(CF)和新宾地区(CX)采集标本。根据形态学测量数据,利用检索表进行物种分类确定[2,14],发现共捕捉到9个物种,对每个物种分别选取1~3个标本进行分子系统学分析,详情见表1。
1.2 实验方法
利用酚-氯仿法对鼩鼱样本的肌肉组织进行基因组DNA提取。mtDNA的Cytb基因全序列PCR反应体系为50 μL,含200 μmol/L的dNTP,1.5 mmol/L的MgCl2,Primer(L14724/ H15915)各10 pmol[15],TaqDNA polymerase 2.5 U(大连宝生物),模板为250 ng。扩增程序为94℃/3 min,(94℃/45 s,50℃/50 s,72℃/45 s)30 cycles,72℃/7 min,4℃/∞。PCR扩增产物经纯化回收,进行测序。
1.3 数据处理
通过GenBank下载与采集标本相对应鼩鼱属9个物种的Cytb基因全序列,以及下载GenBank仅有的鼩鼱属其他30个物种的Cytb基因全序列(表1)。序列使用EditSeq(DNASTAR软件包)进行序列编辑,再利用Clustal W软件进行序列聚类比对。利用MEGA 5.05计算Kimura-2参数校正的遗传距离,及用邻接法(Neighbor-joining,NJ)构建带有Boot-strap检测值的分子系统树。利用分析软件Paup 4.0用最大似然法(Maximum likelihood,ML)构建带有Boot-strap检测值的分子系统树,Boot-strap 1 000次重复检测分子系统树的置信度。利用大麝鼩(Crociduralasiura)做外群。
表1 鼩鼱属分子系统学分析的样本
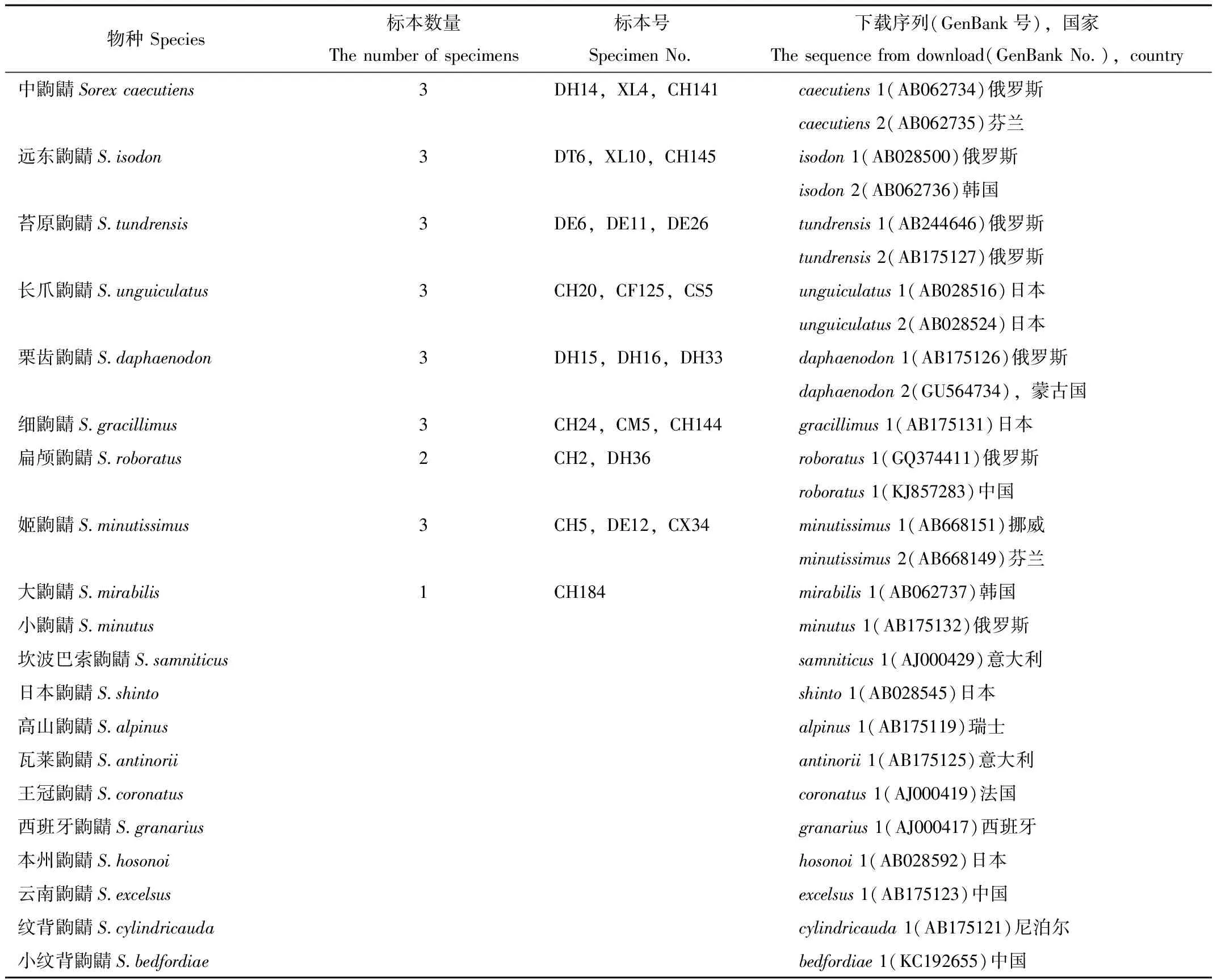
Tab.1 The sample table of molecular phylogenetic analysis on Sorex

续表1
2 结果
2.1 Cyt b 基因全序列的组成与变异
本研究扩增的9种鼩鼱属物种和从GenBank下载与采集标本相对应物种的Cytb基因全序列,连同GenBank下载30个鼩鼱属物种的Cytb基因全序列,共对81个样本序列进行分析。Cytb基因全序列大小均为1 140 bp,其中A、T、G和C的平均含量分别为29.0%、29.0%、13.6% 和28.4%。A+T含量较高,为58.0%,而G+C含量为42.0%。有138个位点发生碱基替换,转换数与颠换数的比值(R)为2.96。
2.2 遗传距离
利用MEGA 5.05计算鼩鼱属39个物种的81个样本的遗传距离。种内遗传距离的范围是0~3.2%,最大种内遗传距离发生在细鼩鼱的种内样本间。种间遗传距离的范围是0.2%~17.3%,最小的种间的遗传距离发生在贫瘠地面鼩鼱(S.ugyunak)和弗兰格尔鼩鼱(S.portenkoi)之间,最大的种间的遗传距离发生在大鼩鼱和特氏鼩鼱(S.trowbridgii)之间,涉及我国东北分布鼩鼱物种的最小的遗传距离(0.6%)发生在阿拉斯加小鼩鼱和姬鼩鼱之间。我国东北地区分布9个鼩鼱属物种的遗传距离见表2,其种间遗传距离的范围是4.8%~11.9%,最小的种间的遗传距离发生在苔原鼩鼱和栗齿鼩鼱之间,最大的种间的遗传距离,发生在中鼩鼱和大鼩鼱之间。
表2 我国东北地区分布的9个鼩鼱属物种的遗传距离
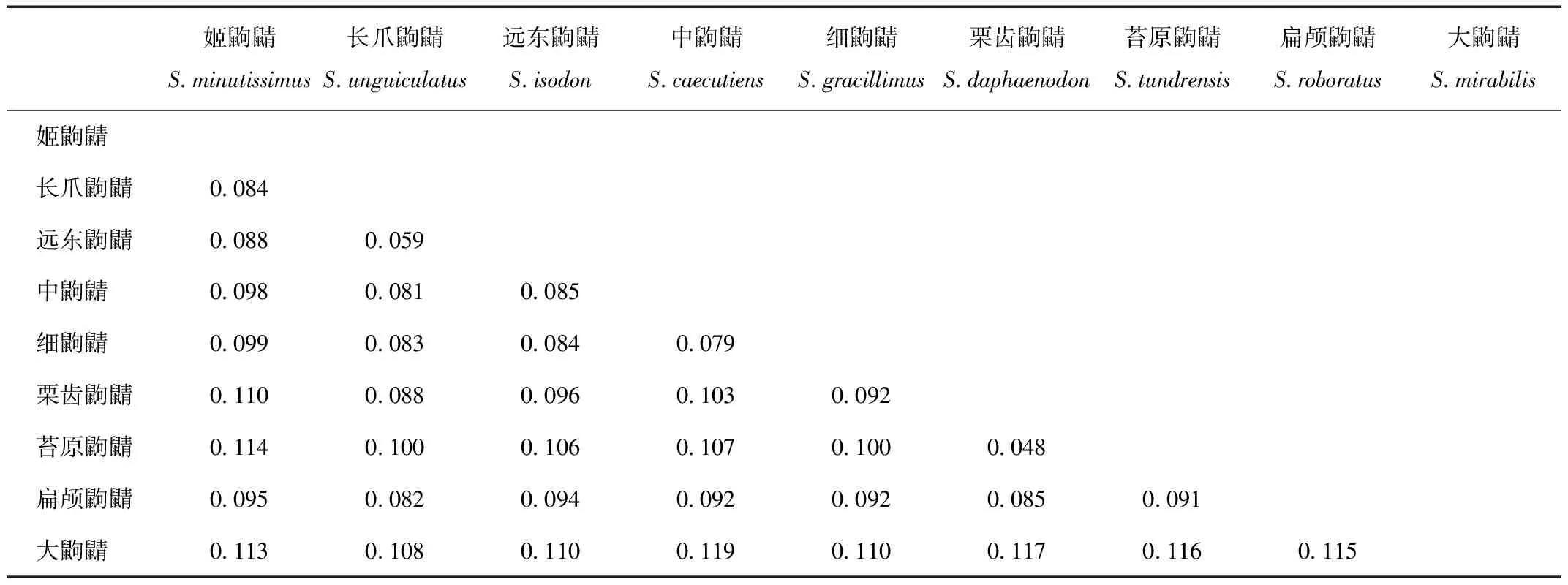
Tab.2 The K2P genetic distance among 9 species of Sorex from Northeast China
2.3 系统发育重建
基于Cytb基因全序列数据,分别利用分析软件MEGA 5.05采用NJ法和利用分析软件Paup 4.0采用 ML法构建分子系统树(图1,图2)。NJ树和ML树表现鼩鼱属物种的系统发生关系是基本一致的。系统发生树首先分成2大分支。一大支基本由分布在古北区的鼩鼱属物种构成,其中阿拉斯加小鼩鼱和北极鼩鼱(S.arcticus)是分布在新北区物种,苔原鼩鼱是分布在全北区的物种。另一大分支基本由分布在新北区的鼩鼱属物种和分布在古北区的堪察加鼩鼱(S.camtschatica)和弗兰格尔鼩鼱构成。
古北区大分支首先分化出来的是高山鼩鼱(S.alpinus),表现出高山鼩鼱较为古老。古北区大分支分化出6个置信度较高(bootstrap value≧63)的分支:中鼩鼱-日本鼩鼱(S.shinto)分支;小鼩鼱-沃氏鼩鼱(S.volnuchini);长爪鼩鼱-远东鼩鼱分支;姬鼩鼱-本州鼩鼱(S.hosonoi)分支,分支内包括阿拉斯加小鼩鼱(S.yukonicus);XY1Y2性染色体复杂群体(Sorexaraneus-arcticusgroup)[16-17],性染色体复杂群体分支包括苔原鼩鼱、天山鼩鼱(S.asper)、普通鼩鼱(S.araneus)、西班牙鼩鼱(S.granarius)、高加索鼩鼱(S.satunini)、瓦莱鼩鼱(S.antinorii)、王冠鼩鼱(S.coronatus)和栗齿鼩鼱;云南鼩鼱(S.excelsus)-纹背鼩鼱(S.cylindricauda)-小纹背鼩鼱(S.bedfordiae)分支,是由我国西南地区具有分布的3个物种组成。在NJ树和ML树都以高置信度支持坎波巴索鼩鼱(S.samniticus)与性染色体复杂群体分支具有较近的系统发生关系。在NJ树和ML树都显示扁颅鼩鼱和杜氏鼩鼱(S.raddei)依次与性染色体复杂群体分支具有一定的系统发生关系,具有较低的置信度,可见这两个物种较为特化。在NJ树和ML树都显示大鼩鼱与云南鼩鼱-纹背鼩鼱-小纹背鼩鼱分支具有一定的系统发生关系,具有较低的置信度,可见该物种也较为特化。细鼩鼱的系统发生地位在NJ树和ML树存在不同,尽管都具有较低的置信度,NJ树显示细鼩鼱与中鼩鼱-日本鼩鼱分支具有较近的系统发生关系,而ML树显示细鼩鼱与姬鼩鼱-本州鼩鼱分支具有较近的系统发生关系。
无论NJ树还是ML树,新北区大分支都以高置信度(≧91%)分成2个分支,其中一个分支是特氏鼩鼱-索绪尔鼩鼱(S.saussurei)分支,这是个较为特化的分支,2个物种彼此之间也以较高置信度分化,说明2个物种也存在一定的系统分化。另个分支是由其他物种构成,尽管由烟熏鼩鼱(S.fumeus)和阴阳鼩鼱(S.tenellus)构成的小分支的系统发生地位,在NJ树和ML树存在不同,但整体这个分支分为4个小分支。由太平洋水鼩鼱(S.bendirii)、沼泽鼩鼱、贝尔德鼩鼱(S.bairdi)、暗黑鼩鼱(S.monticolus)、装饰鼩鼱(S.ornatus)和流浪鼩鼱(S.vagrans)构成了第二小分支。矮小鼩鼱(S.hoyi)和奥林匹克鼩鼱(S.rohweri)为第三个小分支。假面鼩鼱(S.cinereus)群体构成了第四个小分支,尽管这个小分支包括了堪察加鼩鼱和弗兰格尔鼩鼱2个古北区物种,但这个群内物种之间表现出十分近的亲缘关系。

图1 基于Cyt b 基因构建的鼩鼱属NJ系统进化树Fig.1 Phylogenetic tree of Sorex inferred from NJ based on the Cyt b gene

图2 基于Cyt b 基因构建的鼩鼱属ML系统进化树Fig.2 Phylogenetic tree of Sorex inferred from ML based on the Cyt b gene
3 讨论
3.1 鼩鼱属的系统分化
鼩鼱属的亚属分化问题一直存在争议,基于形态学研究认为应将鼩鼱属分为3个亚属:Sorex、Microsorex和Otisorex[18];基于核型分析认为鼩鼱属应分为3个亚属,一个是XY1Y2性染色体复杂群体(Sorexaraneus-arcticusgroup),另一个为新北区群(Otisorex),第三个是除去性染色体复杂外的古北区群体(Homalurus)[19];基于同工酶分析和免疫学分析认为鼩鼱属应分为3个亚属,一个是分布于古北区的亚属(Sorex),另一个为分布于新北区的亚属,第三个是包括特氏鼩鼱在内的未命名亚属[20]。本研究系统进化树明显分为2大分支,进一步支持分布于古北区的亚属和分布于新北区的亚属的分化,这两个亚属的分化与之前的研究还是较为一致的。
一大支基本由分布在古北区的鼩鼱属物种构成,其中阿拉斯加小鼩鼱和北极鼩鼱是分布在新北区物种,苔原鼩鼱是分布在全北区的物种。阿拉斯加小鼩鼱表现出与姬鼩鼱极其近的亲缘关系,阿拉斯加小鼩鼱应该是姬鼩鼱分布在新北区的同种异名体,这与之前基于Cytb基因部分序列和控制区序列针对这2个类群研究结果是一致的[21]。鼩鼱属的系统分化关系与第四纪冰期白令地区在冰期与间冰期表现出的周期性关闭有着明显关系[6]。这样看来姬鼩鼱和苔原鼩鼱是全北区物种,自然它们的系统发生地位就很好理解了,然而北极鼩鼱到目前还没有找到在古北区的分布,推测古北区东北亚应该存在与其亲缘关系很近的物种存在。XY1Y2性染色体复杂群体(Sorexaraneus-arcticusgroup)包括的8个物种[19],明显聚在一个分支上,与基于核型分析结果较为一致,但其分化不足以成为一个亚属,而是在分布于古北区的亚属内部。
另一大分支基本由分布在新北区的鼩鼱属物种和分布在古北区的堪察加鼩鼱和弗兰格尔鼩鼱构成。其中特氏鼩鼱-索绪尔鼩鼱分支,表现出高度特化,这与同工酶分析和免疫学分析是一致的[20],既然是分布于新北区的亚属内的一个高度特化分支,说明本研究认为特氏鼩鼱-索绪尔鼩鼱类群与新北区的亚属具有更近的系统进化关系,与免疫学分析是一致的,不同于同工酶分析认为该类群与古北区的亚属具有更近的系统进化关系[20]。由太平洋水鼩鼱、沼泽鼩鼱、贝尔德鼩鼱、暗黑鼩鼱、装饰鼩鼱和流浪鼩鼱构成的分支内部的系统发生地位与利用染色体和控制区序列分析结果较为一致[10]。烟熏鼩鼱和阴阳鼩鼱小分支和矮小鼩鼱-奥林匹克鼩鼱小分支,在本研究构建的系统进化树中,分化置信度较低,其在新北区的亚属内的系统地位没有得到很好地解决。本研究显示假面鼩鼱群体是个很特别的群体,这个群体在本研究构建的系统进化树中,也高置信度分化成2个小分支,2个小分支内部物种表现出较近的亲缘关系,尤其包括了堪察加鼩鼱和弗兰格尔鼩鼱2个古北区物种的小分支,已经达到了同物种分化水平,因此堪察加鼩鼱和弗兰格尔鼩鼱2个古北区物种的系统分化地位也就很好理解了。
3.2 东北地区鼩鼱属物种的系统发生地位
我国东北分布的9个物种完全聚在古北区的亚属内部,与地理分布是完全相符的。我国东北分布的中鼩鼱与分布于日本的日本鼩鼱聚为一个分支,日本鼩鼱曾作为中鼩鼱的亚种,且两者的核型分析结果是很相似的(2n=42)[6],本研究的分子系统进化树进一步证实了2个物种的分化,且具有很近的亲缘关系。我国东北分布的长爪鼩鼱与远东鼩鼱聚为一个分支,曾经认为长爪鼩鼱是远东鼩鼱的亚种[14],并且两者的核型分析结果是很相似的(2n=42;NF=68)[6],本研究的分子系统进化树进一步证实了2个物种的分化,且具有很近的亲缘关系。我国东北分布的姬鼩鼱与分布于日本的本州鼩鼱聚为一个分支,分支内包括阿拉斯加小鼩鼱(S.yukonicus),像前面分析的那样,本研究支持阿拉斯加小鼩鼱是姬鼩鼱分布在新北区的同种异名体,系统进化树证实了姬鼩鼱和本州鼩鼱的分化,且具有很近的亲缘关系。我国东北分布的苔原鼩鼱和栗齿鼩鼱都聚在XY1Y2性染色体复杂群体(Sorexaraneus-arcticusgroup),这2个物种的确是性染色体复杂物种[16-17],苔原鼩鼱与我国天山有分布的天山鼩鼱表现出很近的亲缘关系,进一步支持了曾经认为两者是姊妹种的研究结果[11,22]。我国东北分布的扁颅鼩鼱在系统发生地位上较为特化,尽管表现出了与性染色体复杂群体分支具有一定的系统发生关系,但其不属于性染色体复杂物种,并且与鼩鼱属其他物种存在较大的分化。我国东北分布的大鼩鼱是东北分布的鼩鼱属物种中最特化的一个物种,表现为我国西南地区具有分布的云南鼩鼱-纹背鼩鼱-小纹背鼩鼱分支具有一定的系统发生关系,但与其他分布于东北地区的物种亲缘关系较远。细鼩鼱的系统发生地位在NJ树和ML树存在不同,尽管都具有较低的置信度,NJ树显示细鼩鼱与中鼩鼱-日本鼩鼱分支具有较近的系统发生关系,而ML树显示细鼩鼱与姬鼩鼱-本州鼩鼱分支较近的系统发生关系。由于细鼩鼱和中鼩鼱都具有颅骨近圆形以及单尖齿整体形态表现为排列紧密,高度较低,每个单尖齿宽度大于高度的共同特征,基于形态学研究认为两者有较近的亲缘关系[23]。最大的种内遗传距离(3.2%)发生在我国东北分布的细鼩鼱和日本北海道分布的细鼩鼱(AB175131)之间,从遗传距离上看该分化是亚种分化,我国东北分布的细鼩鼱和日本北海道分布的细鼩鼱应该是不同亚种。
