荒漠绿洲过渡带柽柳种群结构与空间格局动态
2019-02-21康佳鹏马盈盈马淑琴薛正伟杨丽丽柳维扬
康佳鹏,马盈盈,马淑琴,薛正伟,杨丽丽,韩 路,柳维扬
塔里木大学植物科学学院, 阿拉尔 843300
种群结构是种群最基本的特征,它既可反映种群内不同大小个体数量的分布和配置状况,也能反映种群生存状况与受干扰状态及预测未来种群动态、演变趋势[1-2]。种群空间分布格局是指种群个体在水平空间的配置状况,反映环境对个体行为,生长和生存的影响及相互关系[2-3]。植物种群空间格局受自身生物学特性、种间关系、生境条件及其相互作用的共同影响[3],分析其特征能揭示种群生存策略、空间格局形成的生态学过程、成因及动态机理[2-4]。因此,研究种群结构和空间分布格局,有助于更好地认识种群生态特性、影响因素、形成机制与演替规律,揭示种群和环境间的关系及其对环境资源的利用与适应策略,对种群及群落更新复壮、生态系统恢复具有重要意义。
荒漠绿洲过渡带是干旱区荒漠生态系统的组成部分,其对环境变化敏感且直接影响干旱绿洲区人类生存、绿洲生态系统的稳定和生态安全[5]。多枝柽柳(Tamarixramosissima)作为西北地区优良的防风固沙、盐碱地治理的灌木树种[3],广泛分布于塔里木极端干旱荒漠区,成为维持干旱荒漠生态系统稳定的关键种[4],对遏制区域生态退化、改善生态环境和维护荒漠绿洲生态系统健康发挥着重要作用。多枝柽柳在塔克拉玛干沙漠北缘荒漠过渡带多形成大面积连片的单优群落,也常与其他荒漠植物共存,成为绿洲前沿风沙区的典型代表群落和天然屏障。近年来,随着塔里木灌区人工绿洲的不断扩张,绿洲外围过渡区域遭到强烈挤压,致使柽柳灌丛生长衰退,林分结构失调,系统功能退化。荒漠绿洲生态环境恶化作为一个全球性问题己受到国内外广泛关注和高度重视。目前,学者对干旱荒漠区柽柳灌丛的研究主要集中在水盐环境梯度下柽柳生长发育、群落结构与分布格局、生态适应性、生物多样性、土壤养分空间异质性及种群与环境关系[6-11]等方面,但有关塔克拉玛干沙漠北缘荒漠绿洲过渡带柽柳灌丛结构与空间格局动态及退化机理的研究少见报道。柽柳种群在塔里木荒漠绿洲过渡带的生长状况和种群动态如何?生活史特征是什么?生物因素和环境因素如何影响柽柳种群结构与空间分布格局? 主要限制因子是什么?本文以塔克拉玛干沙漠北缘荒漠绿洲过渡带柽柳灌丛为研究对象,通过野外样地调查分析柽柳种群径级/高度结构、数量与空间格局动态,揭示柽柳种群动态规律与未来发展趋势及其影响机制、退化机理,以期为塔里木极端干旱荒漠区柽柳种群保护、复壮更新与荒漠植被恢复、生态治理提供科学理论依据。
1 研究区概况
研究区位于塔克拉玛干沙漠北缘,阿拉尔市南口镇荒漠绿洲过渡带(81°24′E,40°28′N),海拔1008 m,属暖温带大陆性干旱气候,区内光热资源丰富,昼夜温差大。太阳总辐射能5796 MJ m-2a-1,平均日照时数2729.0 h,年平均气温10.4℃,≥10℃年积温4340℃,平均气温≥10℃的持续日数为201 d,极端最高温度39.4℃,极端最低温度-25.0℃。年平均降水量50.4 mm,年平均蒸发量1880.0 mm,干燥度12—19。风沙灾害频繁,春、夏季多大风天气,是该地区风沙危害的主要季节。过渡带物种稀少,建群种为多枝柽柳(Tamarixramosissima),伴生种有芦苇(Phragmitesaustralis)、骆驼刺(Alhagisparsifolia)、花花柴(Kareliniacaspica)等。地下水位2.5 m左右,土壤类型为风沙土,土壤表层总盐5.36%,质量含水量5.86%,有机质含量1.51 g/kg,全氮含量0.03 g/kg。
2 研究方法
2.1 样地设置与野外调查
经充分踏查在塔克拉玛干沙漠北缘荒漠绿洲过渡带典型代表性地段设置1块100 m×100 m样地,以50 m间隔分成4个50 m×50 m的小样方。采用相邻格子法对天然柽柳灌丛调查,以10 m×10 m为基本单元,记录每个单元内柽柳灌丛数量、丛径数、高度、冠幅、基径等指标,同时以样地一边为X轴,以其垂直边为Y轴建立平面直角坐标系,测量所有柽柳灌丛(X,Y)座标。草本层(1 m×1 m)采用对角线法进行植物种类、高度、频度、多度等详细调查,并用GPS定位,记录样地经纬度和海拔等生态因子。
100 m×100 m样地中心“十”字线上与每块小样方对角线上(5点)采集土样,深度1.0 m,分5层用土钻每隔20 cm取土样分层混合,用铝盒和塑料袋封装,室内测定土壤理化性质。土壤含水量采用烘干法测定;土壤总盐采用蒸馏水浸提,水土比为5∶1,水浴蒸干法测定。土壤有机质采用重铬酸钾-硫酸外加热法测定;全氮采用凯式定氮法测定;速效氮采用碱解扩散法测定;速效磷采用碳酸氢钠浸提-钼蓝比色法测定;全钾采用氢氧化钠熔融-火焰光度法测定;速效钾采用醋酸铵浸提-火焰光度法测定。
2.2 数据分析
2.2.1 柽柳种群结构和生命表、存活曲线
根据柽柳生长缓慢的生物学特性,正常条件下基径大小与灌木年龄成正比。本研究采用基径(BD)和树高(H) 2个指标进行种群结构分析。根据基径(BD)的分布状况,将基径划分为11个等级:Ⅰ级(BD<1 cm)、Ⅱ级(1 cm≤BD<2 cm)、Ⅲ级(2 cm≤BD<3 cm)、Ⅳ级(3 cm≤BD<4 cm)、Ⅴ级(4 cm≤BD<5 cm)、Ⅵ级(5 cm≤BD<6 cm)、Ⅶ级(6 cm≤BD<7 cm)、Ⅷ级(7 cm≤BD<8 cm)、Ⅸ级(8 cm≤BD<9 cm)、Ⅹ级(9 cm≤BD<10 cm)、Ⅺ级(BD≥10 cm)。高度以0.5 m为级阶,每增加0.5 m为一级,将高度划分为9个等级:Ⅰ级(H<0.5 cm)、Ⅱ级(0.5 cm≤BD<1.0 cm)、Ⅲ级(1.0 cm≤BD<1.5 cm)、Ⅳ级(1.5 cm≤BD<2.0 cm)、Ⅴ级(2.0 cm≤BD<2.5 cm)、Ⅵ级(2.5 cm≤BD<3.0 cm)、Ⅶ级(3.0 cm≤BD<3.5 cm)、Ⅷ级(3.5 cm≤BD<4.0 cm)、Ⅸ级(BD≥4.0 cm)。统计样地内各径级/高度级株数,以径级/高度级为纵轴,株数/hm2为横轴,绘制柽柳种群径级/高度结构图。将径级从小到大的顺序看作是时间顺序关系,统计各径级株数,采用均滑技术处理,编制种群静态生命表[12],并绘制存活曲线。
依据柽柳生活史特点,将其划分为4个生长阶段:幼苗(Ⅰ级)、小径级灌木(Ⅱ—Ⅲ级)、中等径级灌木(Ⅳ—Ⅶ级)、大径级灌木(Ⅷ—Ⅺ级)。
2.2.2 柽柳种群结构动态与趋势预测
运用高度级间的失稳率[13],时间序列预测模型[14]对种群数量动态分析。
两相邻高度级(i→i+1级)间种群个体数量失去稳定的速率:
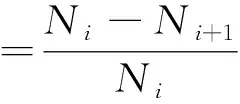
(1)
种群整体失去稳定的速率由各级失稳率V(i,i+1)对各级该种群个体数Ni加权:

(2)
式中,Vi,i+1为相邻高度级失稳率;V(%)为种群整体高度级失稳率;Ni为第i高度级种群的个体数。
时间序列预测模型:
(3)
式中,n表示需要预测的未来时间年限,Mt表示未来n年时t龄级种群大小,Xk为当前k龄级的种群大小。根据现有柽柳种群的年龄结构数据,应用上式计算、预测未来种群发展趋势,原理与方法见文献[14]。
2.2.3 种群空间分布格局与空间关联性
采用扩散系数、丛生指标、聚块性指数、负二项参数、平均拥挤度及Morisita指数等6种指标,计算不同径级柽柳的空间分布类型,具体计算方法见文献[2,15]。为了进一步解释实际的空间格局,建立视植物为二维空间点的 Ripley′sK函数以样地内某点为圆心,一定长度t为半径的圆内植物个体数进行计算,计算公式为[16-17]:

(4)
(5)
式中,K(t)为从某一随意点起,个体落在半径t内的期望值,A为样方面积,n为样方内种群个体数或不同生长阶段的个体数;It(uij)为指示函数,设uij为点i到点j的距离,当uij≤t时,It(uij)=1,当uij>t时,It(uij)=0;Wij为以点i为圆心,uij为半径的圆周长在面积A中的比例,可校正边界效应引起的误差。
应用Ripley′s双变量K12(t)函数对柽柳不同生长阶段种内关联性进行分析,其计算及原理与Ripley′sK函数类似,可用下式估算:
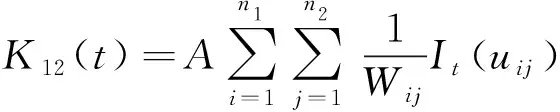
(6)
(7)
式中,K12为从某一随意点起,2个类型个体落在半径t内的期望值;n1、n2为2个类型组的数目;A、It(uij)和Wij含义同上。
为了评价L(t)、L12(t)偏离随机分布的显著性程度,利用Monte Carlo方法通过1000次随机模拟来计算99%的置信区间(上下包迹线)。当L(t)、L12(t)位于上包迹线之上时分别为聚集分布、空间正关联,位于上下包迹线之间时分别为随机分布、相互独立,位于下包迹线之下时分别为均匀分布、空间负关联。采用空间尺度由0增加到50 m,步长为1 m;利用生态学软件包ADE-4来完成。
当种群表现为聚集分布时,把偏离随机置信区间的最大值定义为聚集强度,对应于聚集强度的尺度为聚集尺度,把以对应于聚集强度的聚集尺度为半径的圆面积定义为最大聚集规模[18]。
2.2.4 种群分布与环境的关系
为分析柽柳种群分布与环境因子之间的关系,将土壤水分、总盐、有机质、氮、磷、钾等因子作为自变量,分别与每个小样方内的柽柳个体数量进行相关分析,并对相关系数r的可信度进行t检验[19]。同时采用逐步回归分析得出最优回归方程,确定显著影响因素。
3 结果与分析
3.1 荒漠绿洲过渡带柽柳种群结构动态分析
3.1.1 柽柳种群结构
图1可见,塔里木荒漠绿洲过渡带柽柳种群的个体数量随径级的增大呈先增加后降低的单峰型变化趋势,径级结构呈中间大、两头小的纺锤型,属于衰退型种群,但种群结构保持相对完整。据统计,幼苗(Ⅰ)数量最少,仅3株,占总体1.60%;小径级灌木(Ⅱ—Ⅲ)数量最多共109株,占总体59.98%;中等径级灌木(Ⅳ—Ⅶ)个体数量较多为69株,占36.70%;大径级灌木(Ⅷ—Ⅺ)个体数量较少,有7株,占总体3.72%,表明过渡带柽柳种群幼苗更新较差,随时间推移和灌木继续生长,中等径级、大径级灌木逐渐衰老死亡, 幼苗数量无法及时更新补充,将导致柽柳种群衰败。
柽柳种群高度结构与径级结构变化相似,呈正态分布(图1)。高度(H)<0.5 m的幼苗(Ⅰ)和H<1.0 m灌木(Ⅱ)个体数量最少,分别占总体1.06%、2.13%,仅有6株;Ⅲ—Ⅵ级个体数量最多(163株),占总体86.70%,其中Ⅳ、Ⅴ级个体数量较多。H>3.0 m的大个体(Ⅶ—Ⅸ)数量较小,占总体10.11%。表明过渡带立地条件较差,柽柳幼苗、低矮灌木生长明显受抑,种群更新困难。经统计分析表明,柽柳基径与高度呈对数关系,y=1.3833ln(x)+0.7589,R2=0.9810(n=155,P<0.01),拟合方程达极显著水平。对数方程显示,柽柳种群具有前期生长快,后期生长慢的特征。
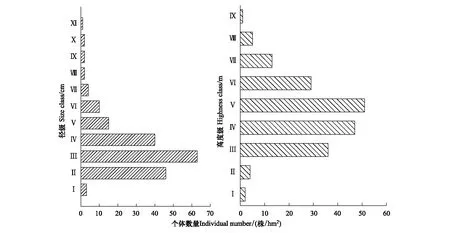
图1 荒漠绿洲过渡带柽柳种群径级与高度结构Fig.1 The size and highness class of T. ramosissima population at the desert-oasis ecotone

图2 柽柳种群存活曲线(Sc)、死亡率(qx)及消失率(Kx)曲线Fig.2 The survival (Sc), mortality (qx) and hazard rate (Kx) curve of T. ramosissima population
3.1.2 柽柳种群静态生命表、存活曲线、死亡率曲线与消失率曲线
表1可见,塔里木荒漠绿洲过渡带柽柳种群存活数随着龄级增加而降低,死亡率在Ⅵ龄达最高,之后随径级增加而下降,呈现“低-高-低”纺锤型模式。原因是随柽柳灌丛逐渐长大,对空间资源需求不断增加与生态位发生重叠,种内个体之间竞争加大,自疏作用加强,导致死亡率不断上升。随径级增大与冠幅扩大,种内竞争更加激烈,导致死亡率出现峰值,其后个体数量减少、竞争减弱、死亡率下降。期望寿命(ex)随径级增大而逐渐下降(表1),是由于小径级个体间竞争较弱,能有效地利用群落资源,从而种群进入生长旺盛期,ex较大,即年轻个体生命力与平均生存能力强。随后ex在Ⅶ、Ⅷ径级有所回升,然后又下降,主要是柽柳种群达到一定年龄后生理机能开始退化,这种趋势符合种群的生物学特征,反映了柽柳种群整个生命过程中个体生命力与平均生存能力的衰退。
柽柳种群存活曲线(Sc)表明幼苗、小径级灌木存活率较高,中等径级灌木(Ⅳ—Ⅶ)存活率较低而死亡率显著升高(图2)。因为前期个体较小,对空间资源竞争较弱;随径级增长灌丛扩大,对空间资源需求增大、自疏作用增强及环境筛选择强度增大,导致死亡率增高,存活个体减少。Ⅶ径级之后的大径级灌木存活数下降减缓、变化趋于平衡。
柽柳种群死亡率(qx)曲线和消失率(Kx)曲线大致重合,变化趋势一致(图2)。qx和Kx都出现3个高峰期。第一、二高峰期均出现在Ⅳ、Ⅵ径级(中等径级灌木阶段),正是柽柳生长旺盛期,种内资源竞争激烈,生长势较弱的个体死亡,导致其qx、Kx较高。第三高峰期出现在Ⅸ径级,由于此时大径级灌木个体达到一定年龄后生理机能开始衰弱,生存能力降低所导致。采用两种数学模型对柽柳种群存活状况是符合DeeveyII型还是符合DeeveyIII曲线进行检验,经拟合建立其相应模型如下:Nx=8.5523e-0.1067x(r=0.9840**);Nx=8.9158x-0.4284(r=0.8879**)。由于指数模型相关系数(r)大于幂函数模型r值,判定塔里木荒漠绿洲过渡带柽柳种群存活曲线更趋于DeeveyII型。
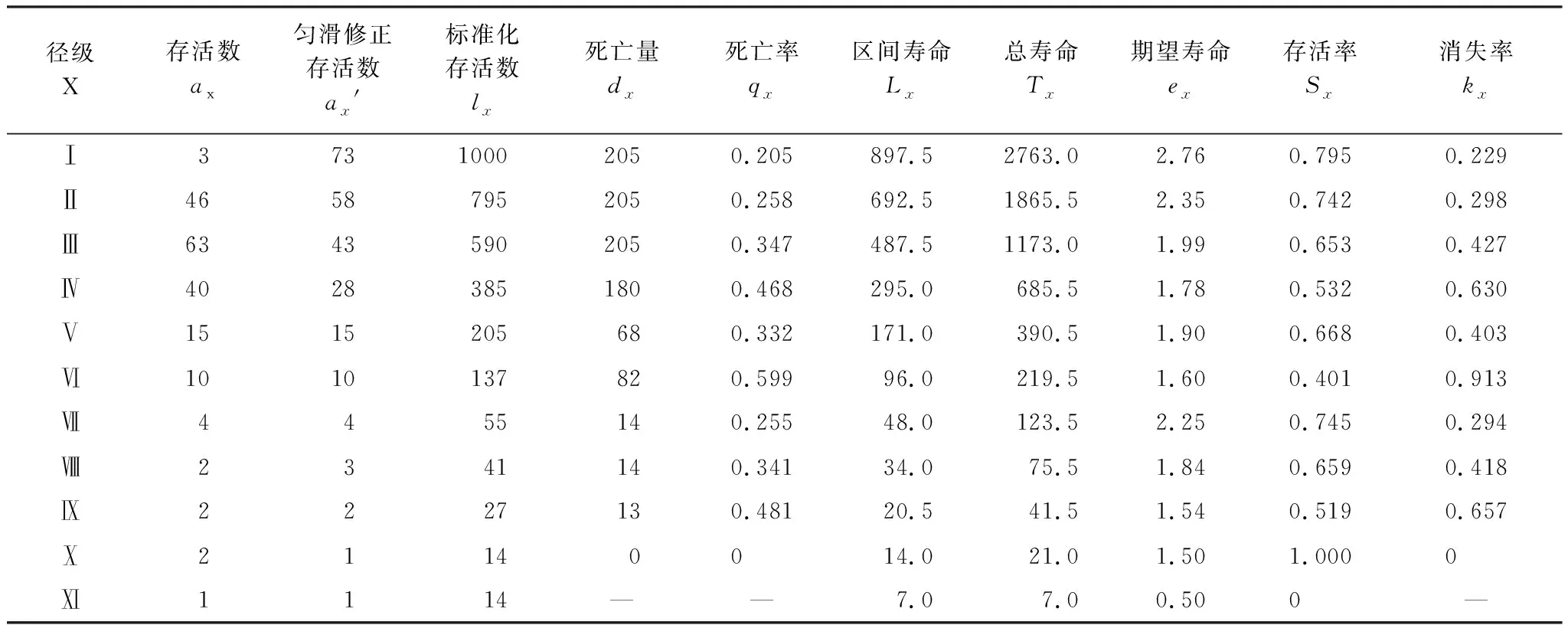
表1 荒漠绿洲过渡带柽柳种群特定时间生命表
X:Size class;ax:Survival numbers;ax′: survival numbers after smoothed;lx:Standard survival numbers;dx:Number of dying during interval;qx:Average mortality rate per age stage;Lx:Lifebetweentheinterval; Tx:Totallife; ex:Meanexpectativelife; Kx:Hazardrate; Sx:survivalrate
3.1.3 柽柳种群高度级失稳率与数量动态预测
利用高度级失稳率判定种群和群落增长、衰退、稳定及量变程度的动态属性[13]。柽柳种群Vp(Ⅰ,Ⅱ)、Vp(Ⅱ,Ⅲ)、Vp(Ⅲ,Ⅳ)、Vp(Ⅳ,Ⅴ)为负值(表2),表明Ⅰ级幼苗,Ⅱ、Ⅲ、Ⅳ级灌木均具有较高的衰退速率,其中Ⅱ级灌木衰退速率最高,其次为Ⅰ级幼苗。Vp(Ⅴ,Ⅵ)、Vp(Ⅵ,Ⅶ)、Vp(Ⅶ,Ⅷ)、Vp(Ⅷ,Ⅸ)均为正值,但并不能说明Ⅴ—Ⅸ级灌木具有较强的增长趋势或较快的增长速率,Vp(Ⅶ,Ⅷ)和Vp(Ⅷ,Ⅸ)正值较高是由于Ⅷ、Ⅸ高度级个体数量较少造成的。过渡带柽柳种群总体失稳率为正值,但非常接近零,表明随着群落演替优势种群正朝着削弱现有组成格局方向发展,种群更新资源匮乏,高度级大的柽柳数量减少与优势地位被削弱,群落朝衰退方向发展。
以塔里木荒漠绿洲过渡带柽柳种群各径级株数为原始数据,按照一次平均推移法预测出各径级在未来3、4、5年后的种群动态(图3)。柽柳种群各径级株数的峰值在时间序列预测中依次向后推移,幼苗(Ⅰ)、小径级灌木(Ⅱ、Ⅲ)个体数随着时间的推移逐渐减少,Ⅴ径级之后各径级个体数逐渐增多。未来3、4年Ⅲ、Ⅳ径级分别比初始种群数量下降了40.74%、5.01%,而未来4年Ⅴ龄级和5年Ⅵ龄级显著增多,其后均呈下降趋势。由于中等径级、大径级灌木数量稳步上升,而幼、小径级株数逐步减少,表明随着时间推移,荒漠绿洲过渡带柽柳种群向老龄化发展、走向衰败。

表2 不同高度级柽柳种群失稳率

图3 荒漠绿洲过渡带柽柳种群数量动态的时间序列预测 Fig.3 Time sequence prediction of number dynamics of T. ramosissima population at the desert-oasis ecotone
3.2 荒漠绿洲过渡带柽柳种群空间分布格局与空间关联性
3.2.1 荒漠绿洲过渡带柽柳种群空间分布格局
柽柳种群总体、幼苗、小径级灌木空间格局均呈聚集分布(表3),中等径级灌木Morisita指数检验(F值)与扩散系数的t检验均不显著,判定其为随机分布;大径级灌木则呈随机分布,表明柽柳种群随生长发育进程其聚集强度逐渐减弱,空间格局由聚集过渡到随机分布。
塔里木荒漠绿洲过渡带柽柳种群空间分布格局随尺度变化明显不同,表现出明显的空间异质性(图4)。点格局分析表明(图5),柽柳种群在<3 m和≥36 m的空间尺度上呈随机分布,其他尺度上均呈聚集分布,且聚集强度随尺度增大逐渐增强,30 m达最大,最大聚集规模为2461.76 m2(图5)。柽柳种群随生长发育其聚集强度逐渐减弱,过渡为随机分布。小径级灌木在<3 m尺度内呈随机分布,其他尺度均呈聚集分布,且聚集强度随尺度增大而增强,41 m达最大(图5)。中等径级灌木仅在<1 m尺度内呈聚集分布,其他尺度均呈随机分布(图5)。大径级灌木在1—13、17—36 m尺度内呈随机分布,而在14—16、>36 m尺度上则呈聚集分布,聚集强度随尺度增大而增强,47 m达最大(图5)。表明荒漠绿洲过渡带柽柳种群随生长发育进程灌丛逐渐长大,个体间对空间资源竞争增强,使种群聚集强度减弱,逐渐过渡为随机分布。
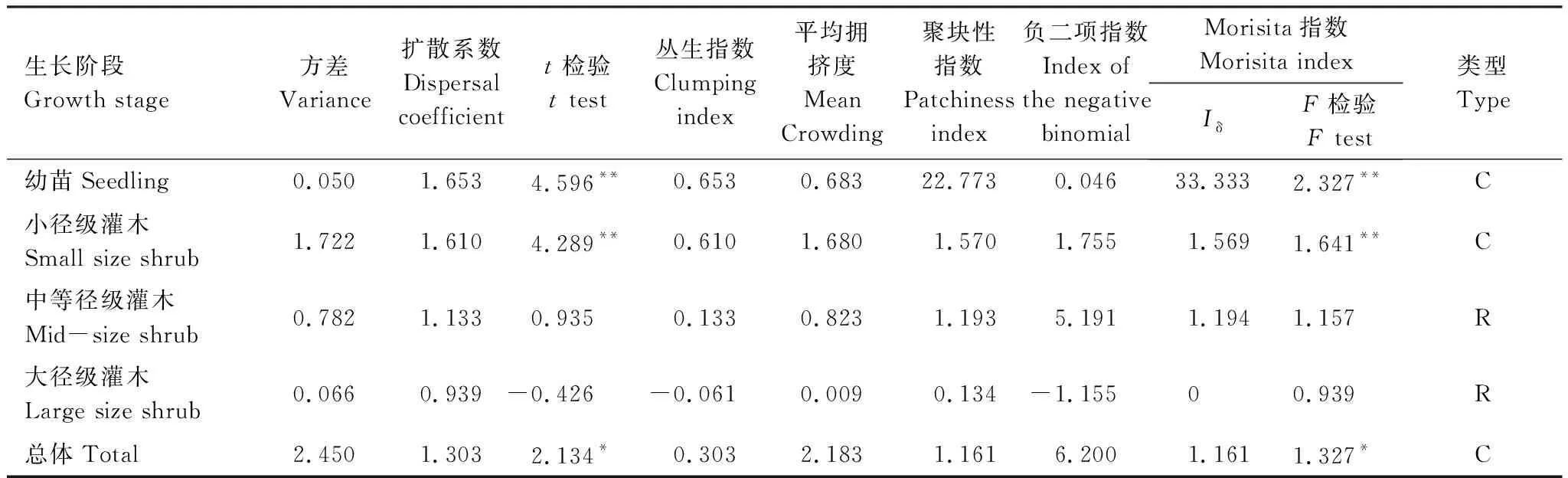
表3 不同生长阶段柽柳种群的空间分布格局
注:C:聚集分布,Clumped distribution,R:随机分布,Random distribution,n=100,*P<0.05,**P<0.01
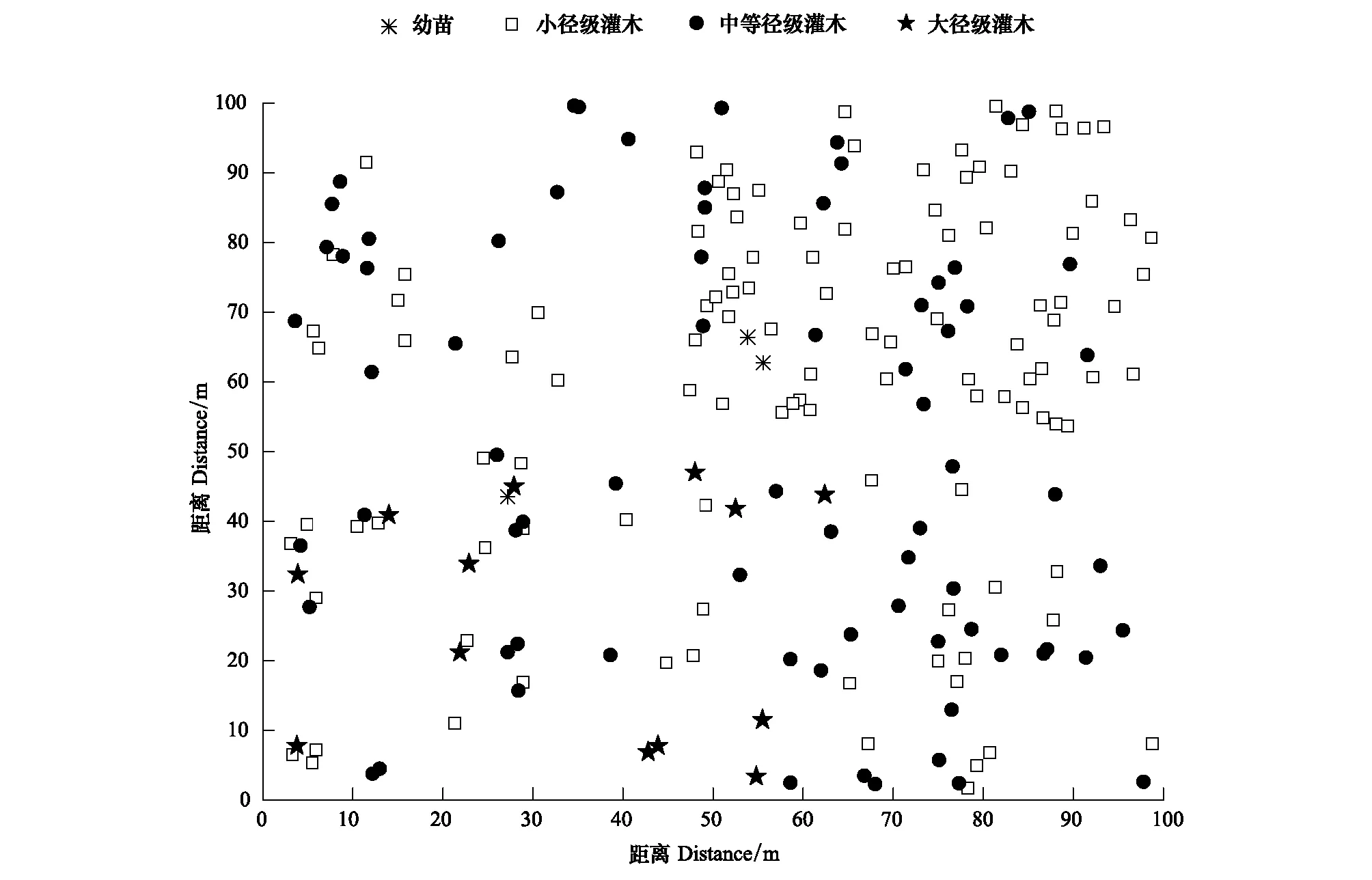
图4 不同生长阶段柽柳空间分布Fig.4 The Point pattern of different growth stages of T. ramosissima population

图5 不同生长阶段柽柳种群点格局分析Fig.5 Point pattern analysis of T. ramosissima population under different growth stages
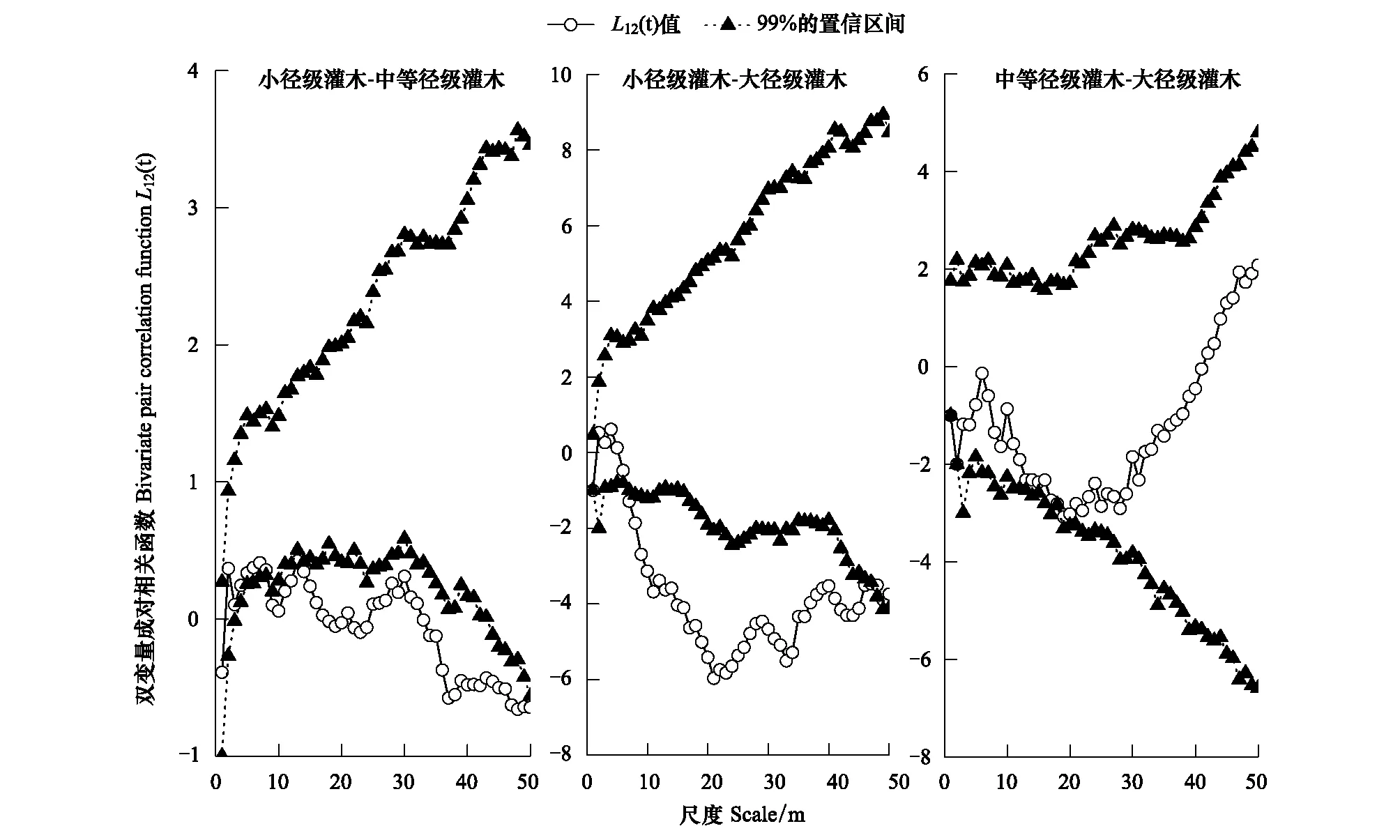
图6 柽柳种群不同生长阶段之间的空间关联性Fig.6 Spatial associations of T. ramosissima population under different growth stages
3.2.2 荒漠绿洲过渡带柽柳种群不同生长阶段间的空间关联性
柽柳种群不同生长阶段之间的RipleyL12(t)双变量空间分析表明(图6),小径级与中等径级灌木、小径级与大径级灌木的空间关联性分别在≥9、7—47 m空间尺度上相互排斥,其他空间尺度上呈相互独立分布,尤其在<7 m尺度范围内个体间呈现非空间依赖性,各自占据自身适宜的生态空间而正常生长。中等径级与大径级灌木的空间关联性在0—50 m空间尺度上均呈相互独立分布,这是因为随个体生长发育,长成中等径级、大径级灌木的个体数量逐渐减少(图1)与空间距离增大、稀疏,使二者在空间尺度上分离而呈独立分布。
3.3 环境与种群分布的相关分析
柽柳种群多度分布与土壤含水量呈显著正相关(P<0.05)(表4),与土壤化学性质均呈负相关,其中与土壤氮素、总盐的负相关系数较高。利用逐步回归分析环境因子与种群多度的关系,得最优回归方程:y=-0.033+0.202x8(水分)+0.013x2(氮素) (P<0.01)。F检验表明,此回归模型可信度达到99.9%(R2=0.998),说明多度变异平方和有99.9%是由土壤水分与氮素变化造成的。因此,土壤水分、氮素是影响荒漠绿洲过渡带柽柳种群生长与分布的主要因子。
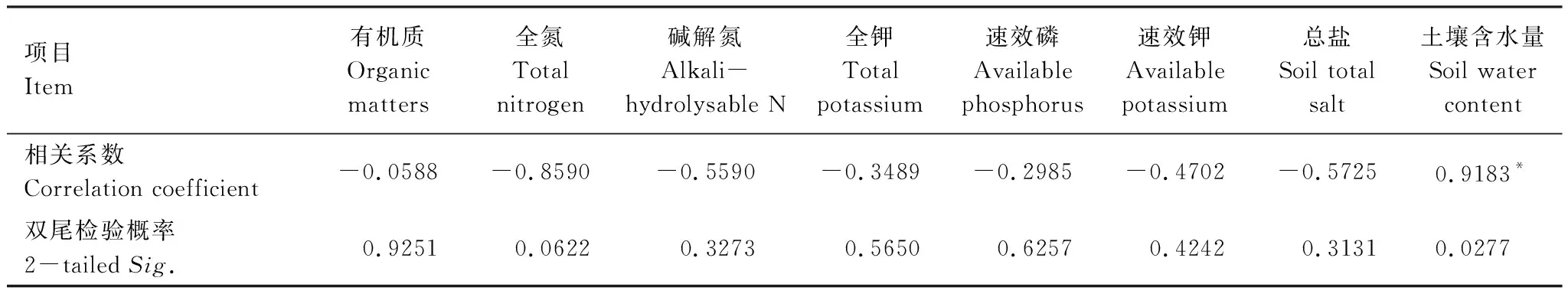
表4 荒漠绿洲过渡带柽柳种群多度与环境因子之间的相关性
n=5, *P<0.05
4 讨论
柽柳是塔里木荒漠绿洲过渡带的生态关键种,其群落生态特征很大程度上影响着极端干旱区生态系统的健康和稳定。加强柽柳种群生态学研究,无论是对荒漠绿洲过渡带植被恢复与生态重建,还是对绿洲生态环境改善、农业生产、社会经济发展都具有重要的意义。
塔里木荒漠绿洲过渡带柽柳群落结构简单、物种组成稀少,在低水高盐生境下大多形成面积较大的单优群落。野外调查发现,柽柳种群径级与高度结构均呈两头小、中间大的纺锤型,更新资源匮乏,属于典型的衰退型种群。表明过渡带生境严酷、强大的环境筛抑制幼苗的建立、存活和生长发育,导致更新困难。学者研究指出柽柳在发芽期和幼苗期常要求湿润的土壤环境,而且一般要求土壤含盐量低于0.15%[20]。绿洲边缘区大量植被被破坏、开垦成农田与农田压碱洗盐退水的排放,在强烈蒸发下林间地表盐分表聚,造成生存环境日益恶化、种子难以萌发和长势衰败。可见,人类不合理资源利用致使柽柳结实困难、种子难以萌发形成幼苗及根萌蘖难以成活,繁殖困难及幼苗无法向幼树阶段转换已成为该种群更新和发展的瓶颈。未来优势种若没有幼苗及时补充,随着演替进行,中等径级、大径级个体不断死亡,数量逐渐减少,种群维持将出现困难。因此,柽柳种群衰退与人为干扰、生境破碎化及生境恶化有关。
荒漠绿洲过渡带柽柳种群存活数、期望寿命(ex)随径级增加而降低,存活曲线接近DeeveyⅡ型,死亡率与消失率变化趋势一致且均出现3个高峰期,分别于Ⅳ、Ⅵ、Ⅸ径级出现高峰。可见,柽柳灌丛逐渐长大为争夺空间资源而导致激烈的种内竞争产生自疏作用及环境筛选择强度增大,是导致中等径级灌木死亡率高的原因;同时伴随着柽柳种群整个生命过程中个体生命力与平均生存能力的衰退。柽柳种群幼苗、低矮灌木高度级失稳率均为负值,尤其Ⅰ、Ⅱ高度级衰退速率最高,虽然种群总体失稳率为正值,但接近零,表明随着群落演替发展,优势种群正朝着削弱现有组成格局方向发展,幼苗无法及时更新和幼龄生长受抑,高度级较大的柽柳优势地位将被逐渐削弱,使过渡带柽柳种群走向衰败。种群动态预测表明, 柽柳种群各径级株数的峰值在时间序列预测中依次向后推移,幼苗、小径级灌木数量大幅减少,中等径级、大径级灌木数量显著增加,柽柳种群将面对死亡高峰。可见,柽柳种群生存动态与其种内对空间资源的激烈竞争、生理功能下降及生境恶化有关。因此,改善生存环境,促进幼苗建立与提高成活率是柽柳种群恢复的关键。
种群空间格局和空间关联性是空间格局分析的两个主要研究内容,是种群生态关系在空间上的两种表现形式[21]。空间分布格局是由种群本身生物学特性、种内种间关系以及环境条件共同影响决定的,其在一定程度上能解释群落结构的形成机理与潜在的生态学过程及环境变化过程,对物种生长、繁殖、死亡、资源利用及对干扰的反应等具有显著的影响[21-22]。种群空间格局与空间尺度密切相关,还受生境异质性和扩散限制的影响。在较小尺度上可能是由种内种间竞争、种子扩散限制、幼苗补充过程等因素所致;在较大尺度上则可能是由物种分布区环境的异质性(如地形、土壤水分等)决定的[22-25]。塔里木荒漠绿洲过渡带柽柳种群分布格局总体为聚集分布,随生长发育进程空间格局从幼苗、小径级灌木的聚集分布过渡至中等径级、大径级灌木的随机分布。径级结构的差异导致种群空间格局上的不同,可能是柽柳在小径级阶段以聚集形式占据空间资源,有利于改善群落微环境,增强种群对不良环境的抗性而存活[26];随生长发育进程植株/灌丛逐渐长大,个体所需空间资源增多、种内竞争加剧,再加上生境逐年恶化与生理功能下降,生长势弱的个体死亡与株间距离扩大,至中等径级、大径级灌木阶段转为随机分布。同时,近年来区域大量垦荒与农田排水使过渡带地下水位上升(2.5 m左右),但极端干旱区地表蒸发与风蚀强烈使土壤浅层水分匮乏与盐分表聚,小径级个体聚集在大径级灌木附近依靠大径级灌木的水力提升作用获得生存水资源[27];随着个体长大和根系下扎,中等径级、大径级灌木对土壤不同层次水源实现分层利用,而空间资源竞争则逐渐增强,自疏作用使种群密度降低,导致不同生长阶段的空间格局发生改变。另外,柽柳不同生长阶段的空间格局也随空间尺度发生变化,小径级灌木在<3 m尺度内呈随机分布,其他尺度均呈聚集分布且聚焦强度随空间尺度增大而增强;中等径级灌木在<1 m尺度内呈聚集分布,其他尺度均呈随机分布;大径级灌木分别在1—13、17—36 m和14—16 m、>36 m尺度内呈随机、聚集分布,这是生境异质性和种内竞争的结果。<3 m的空间尺度生境相对均一,且小径级个体生存需要的资源较少,竞争相对较弱,使其空间格局表现为随机分布;随空间尺度增大,生境异质性增强,尤其土壤水分、养分的分布不均一使其空间格局呈现聚集分布。随径级、尺度的增大,受生境异质性的影响和个体间对光照、水分、养分等有限资源的争夺导致种内竞争加剧, 双重因素作用下使个体数量递减,而减弱种群分布的聚集程度,逐渐形成随机分布。
空间关联性有助于了解群落内物种之间的相互作用及不同物种在不同生境中定居的分异[28],其体现了物种生态幅差异及对生境分化的反应,通常是由于群落生境的差异影响了物种分布所引起[21]。过渡带柽柳种群不同生长阶段之间的空间关联性除小径级与中等径级灌木、小径级与大径级灌木分别在≥9 m、7—47 m尺度呈显著空间负相关,其他尺度上均表现出相互独立分布,表明较大个体为了满足自身的生长发育,不断地与幼小个体争夺有限的空间资源而致使幼小个体生长发育迟滞或死亡(表1),导致种群更新困难,这也间接证实柽柳种群的空间分布是从聚集到随机的自疏过程。因此,荒漠绿洲过渡带柽柳种群不同生长阶段在空间上形成不同的分布格局与空间关联性,与植物群落的自然稀疏过程、干扰格局、资源利用、生境异质性、繁殖特性以及物种对生境的选择有关[18],反映出种群在生长发育过程中的一种生存策略和适应机制。
植被与其生存环境之间是一种相互依赖和制约的关系。塔克拉玛干沙漠北缘荒漠绿洲过渡带降水稀少,蒸发强烈,盐分表聚明显,土地沙漠化与生态退化严重。土壤水盐成为干旱荒漠区诸多生态系统过程的驱动力和关键的非生物限制因子,制约着天然植被的分布、生存和演替[29]。植物个体在适应和生存竞争过程中始终面对由非生物和生物资源所导致的土壤异质性作出响应,而使种群生活史格局发生适应改变。柽柳属植物耐旱、耐盐碱和抗风沙,但过渡带土壤表层盐分含量高(5.36%)且0—60 cm土壤平均含盐量(3.13%)接近其最高耐盐限度(32 g/L)[30]而直接抑制柽柳生长发育;且土壤盐分含量高会降低土壤水势造成柽柳发生生理性干旱。同时,过渡带柽柳密度较低(188丛/hm2),凋落物少,且区域沙尘暴频发和风蚀作用下土壤沙化致使土壤养分大量损失和贫瘠化[31],这些不利因素均直接限制柽柳更新与生长发育。相关与逐步回归分析表明,土壤水分、氮素是限制柽柳种群生长、分布和造成种群更新困难、衰退的主要环境因子。
5 结论
(1) 塔里木荒漠绿洲过渡带柽柳种群结构呈纺锤型,存活曲线接近DeeveyⅡ型,小径级死亡率、消失率与高度级失稳率较高,生境恶化、更新资源匮乏是种群走向衰败的原因。因此,改善生存环境,保护现有植株、促进幼苗建立与提高成活率是柽柳种群恢复的关键。
(2)柽柳种群空间格局总体为聚集分布,随生长发育逐渐从聚集过渡为随机分布;同时分布格局也随空间尺度变化而变化,尤其中大径级对小径级个体生长产生明显抑制作用,直接影响种群更新稳定。种群空间格局变化是生境异质性、种间竞争、更新限制和生态适应的结果。
(3)荒漠绿洲过渡带土壤水分、氮素是限制柽柳种群生长、分布和造成种群更新困难、衰退的主要因子。
