葡萄糖转运蛋白在血糖稳态调节中的功能
2019-02-21张凯艺叶华琼杨述林
张凯艺, 谢 宁, 叶华琼,2, 杨述林*
1.中国农业科学院北京畜牧兽医研究所, 北京 100193;2.安徽科技学院动物科学学院, 安徽 滁州 233100
葡萄糖是人体的主要供能物质,其代谢过程中产生的各种中间产物也可为其他营养物质的代谢提供碳架和前体。机体的血糖水平受到多种因素的精密调节,血糖过低会引起中枢神经系统损伤;血糖过高则会造成多种组织器官大血管和微血管糖毒性伤害。血糖稳态失衡与肥胖、糖尿病、非酒精性脂肪肝等多种代谢性疾病的发生密切相关。国际糖尿病联盟2017年底统计结果显示,全世界糖尿病患者人数已达4.25亿[1],其中以胰岛素抵抗(insulin resistance,IR)并伴有胰岛素分泌不足为特征的T2DM患者约占90%。T2DM的发生受遗传、饮食和生活方式等综合因素影响,病因多样,发病机制复杂,目前仍缺乏有效治疗方案。通过全基因组关联分析,人们已经筛选出100多个T2DM易感基因[2],其中已成功应用于基因诊断和药物开发的基因包括:钾离子通道蛋白(KCNJ11)、胰高血糖素样肽-1受体(GLP-1R)、过氧化物酶体增殖物激活受体γ(PPARγ)等基因[3~5],它们在胰岛素分泌、糖脂代谢和细胞增殖等关键通路中发挥重要作用。
血糖稳态的调控依赖多个组织器官的协同作用,各组织通过吸收、消耗、储存葡萄糖或释放激素及代谢产物来感知或调节血糖[6]。葡萄糖转运蛋白(glucose transporter, GluT)由SLC2A(solute linked carrier family 2, subfamily A)基因编码,包括3组共14个成员,其中Ⅰ组GluT(GluT1、GluT2、GluT3和GluT4)在体内分布最广泛、表达量最高,发挥最主要的葡萄糖转运功能[7, 8]。Ⅰ组GluT成员的底物转运特性与其分布组织在血糖稳态调节中的功能相适应(表1)。GluT1具有与基础血糖相近的Km值,维持全身各组织器官葡萄糖摄取效率的稳定[9];GluT2对葡萄糖具有低亲和力和高转运效率,对高糖环境敏感,有助于肝脏高效发挥血糖调节的功能,并在神经系统的葡萄糖感受中起到关键作用,已成为糖尿病药物研发的重要靶标[10];GluT3的底物亲和力最高,保证中枢神经系统优先利用葡萄糖[11];GluT4是Ⅰ组GluT中唯一胰岛素敏感的转运蛋白,骨骼肌和脂肪的胰岛素敏感性依赖于GluT4的功能;GluT4表达与囊泡转位缺陷与T2DM患者外周组织的IR密切相关,已成为抗T2DM药物开发研究的热点[12]。目前,从机体血糖稳态整体调节角度对Ⅰ组GluT功能及作用的系统介绍较少。本文分别对胰岛、肝脏、脂肪、骨骼肌、神经系统和大脑等关键组织在血糖稳态调节中的作用,以及Ⅰ组GluT在这些组织中的功能进行阐述,以期为T2DM及相关代谢疾病的诊断、治疗和动物模型构建等提供参考。

表1 Ⅰ组葡萄糖转运蛋白的分布与功能Table 1 Distribution and function of Class Ⅰ of GLUTs.
1 GluT在胰岛β细胞中的作用
胰岛在机体血糖稳态调控的过程中发挥核心作用,机体主要的两大血糖调节激素——胰高血糖素和胰岛素,分别由胰岛α细胞和胰岛β细胞合成分泌。胰岛β细胞感受血糖升高并分泌胰岛素的过程被称为葡萄糖刺激的胰岛素分泌(glucose-stimulate insulin secretion,GSIS):餐后血糖上升,经GluT进入β细胞的葡萄糖增加,并在葡萄糖激酶(glucose kinase,GK)的作用下迅速转化为6-磷酸葡萄糖(G6P),其进入线粒体氧化分解,产生大量ATP;ATP/ADP比的升高导致ATP依赖性K+通道关闭,使膜发生去极化,引起电压门控性Ca2+通道开放,Ca2+内流,导致胰岛素的释放(图1)。葡萄糖磷酸化为G6P的过程是GSIS及胰岛素释放的限速步骤,GK基因缺陷会导致青少年患成人型糖尿病(MODY)。虽然葡萄糖转运并不是胰岛素分泌的限速步骤,但GluT与高Km的GK共同构成了β细胞对血糖的感受器,起着允许作用[13],β细胞的葡萄糖转运过程是GSIS必须的。
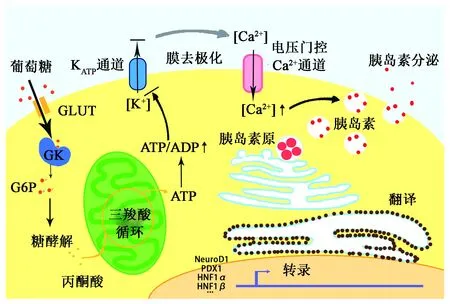
图1 胰岛β细胞GSIS的机制Fig.1 Mechanism of GSIS in islet β-cells.注:GLUT:葡萄糖转运蛋白;GK:葡萄糖激酶;G6P:6-磷酸葡萄糖;PDX-1:胰腺十二指肠同源盒因子1;HNF1α:肝细胞核因子-1α;HNF1β:肝细胞核因子-1β。
值得注意的是,啮齿动物与人类β细胞表面的GluT种类存在差异,啮齿动物β细胞以低底物亲和力的GluT2(Km≈16.2mmol/L)为主要葡萄糖载体,而人类β细胞以较高底物亲和力的GluT1(Km≈6.9mmol/L)为主。GluT2敲除的小鼠缺乏GSIS,并且在10~15日龄即表现出胰岛素分泌障碍、高血糖、血浆胰高血糖素和游离脂肪酸(free fatty acid, FFA)水平上升等T2DM症状,大部分个体在2~3周龄时死亡[14];而在其β细胞恢复表达GluT1或GluT2后,GSIS都可以得到恢复[15];说明β细胞的GSIS依赖于细胞膜上GluT的总体葡萄糖转运能力,而非某一特定类型的GluT。
GluT表达的缺失造成β细胞葡萄糖摄取的障碍,从而失去GSIS[16]。例如:T2DM患者β细胞对于葡萄糖的敏感性降低与其GluT1和GluT2表达下调相关[17,18]。Zuker肥胖糖尿病大鼠、自身免疫性糖尿病大鼠等糖尿病动物胰岛β细胞GluT2表达下降。高脂饮食诱导小鼠的血浆FFA水平上升,GluT2表达下降,并出现肥胖相关糖尿病的早期症状[19];在移植健康鼠的胰岛后,糖尿病小鼠β细胞的GluT2表达和GSIS可以恢复[20,21]。以上研究表明,依赖于GluT的β细胞葡萄糖摄取障碍是糖尿病发生的机制之一。
2 GluT在肝脏中的作用
肝脏是血糖稳态调节的关键组织。正常情况下,肝脏的糖原合成、分解以及糖异生在维持血糖浓度稳定的过程中发挥主要作用。肝脏在餐后血糖升高和胰岛素升高条件下快速摄取葡萄糖并以肝糖原形式储存,而饥饿、运动及其他胰岛素与抗胰岛素因子比例降低时将葡萄糖释放到血液循环。肝脏表面最主要的GluT是GluT2,葡萄糖的流向取决于其在肝细胞内外的浓度差。肝细胞表面的GluT2具有高Km值,因而在餐后高血糖时具有很高的葡萄糖摄取效率。通过GluT2进入肝细胞的葡萄糖在GK的作用下迅速转化为G-6-P,维持细胞内较低的葡萄糖浓度,使血液中的葡萄糖得以持续流入肝细胞并储存。肝细胞中升高的葡萄糖通过磷酸戊糖途径产生的5-磷酸木糖激活蛋白磷酸酶2,使碳水化合物反应元件结合蛋白(ChREBP)去磷酸化[22,23]并进入细胞核,激活丙酮酸激酶、乙酰CoA羧激酶和脂肪酸合酶等基因的转录,从而加强肝细胞的糖酵解和脂合成。同时,胰岛素信号通过Akt通路促进叉头样转录因子O1(FOXO1)的磷酸化,使葡糖异生相关酶表达水平下调,降低糖异生和肝糖输出,并增加肝脏的糖原合成与储存[10]。空腹状态下,血液循环中低血糖、低胰岛素和高胰高血糖素的环境使肝脏葡萄糖激酶活性降低,糖原分解和糖异生相关酶活性升高,导致肝细胞葡萄糖浓度的升高,并由GluT2顺浓度梯度扩散入血窦。
在小鼠肝脏中特异性敲除GluT2(LG2KO)后[24],肝脏的糖摄取受阻,但LG2KO鼠的骨骼肌葡萄糖摄取能力升高,从而代偿性地维持餐后血糖平衡;同时,由于LG2KO鼠缺乏依赖GluT2的肝脏糖输出过程,空腹状态下肝细胞内葡萄糖和G-6-P浓度升高,并通过ChREBP激活糖酵解和脂合成相关基因的表达;另外,LG2KO鼠血液和粪便胆汁酸水平下降,并逐渐表现出β细胞功能障碍和葡萄糖不耐受。以上研究结果说明,GluT2在肝脏糖代谢相关基因的表达调控中发挥了重要的作用,并可以进一步通过胆汁酸-Fxr-KATP途径调节β细胞的GSIS效应[25,26]。虽然肝脏GluT2基因的缺失不会直接影响血糖稳态,但肝脏GluT2缺失引起的葡萄糖代谢的改变可以影响β细胞的功能,最终导致血糖稳态调节障碍。
3 GluT在脂肪中的作用
脂肪作为高容量的能量储存库,在维持机体血糖稳态过程中发挥着重要作用。脂肪组织是最重要的胰岛素敏感组织之一,胰岛素可以促进其成脂分化、葡萄糖摄取以及甘油三酯合成。脂肪也是重要内分泌器官,最早发现的瘦素[14]具有抑制食欲和抵抗肥胖功能;脂肪组织还可分泌脂联素、抵抗素和肿瘤坏死因子α(TNF-α)等细胞因子,在肥胖介导的胰岛素敏感性下降和慢性炎症中起着至关重要的作用。脂代谢紊乱引起的甘油三酯和FFA水平升高是T2DM的典型临床症状。
脂肪组织的胰岛素敏感性依赖于GluT4。无胰岛素作用时,GluT4主要储存在储存囊泡中,并通过循环囊泡实现细胞膜表面与胞内GluT4的动态平衡。胰岛素信号作用于胰岛素受体,通过Akt等通路激活GluT4的囊泡运输,使GluT4迅速大量分布于细胞膜表面,从而激活脂肪的葡萄糖摄取[27]。在胰岛素作用下,虽然进入脂肪组织的葡萄糖只占10%[6],但脂肪特异性敲除GluT4的小鼠出现骨骼肌和肝脏IR、高血糖以及高胰岛素血症,与GluT4全身敲除小鼠相似[28~30]。在骨骼肌敲除GluT4的小鼠的脂肪中特异性过表达GluT4后,小鼠的脂肪沉积和血液FFA水平略有上升,但却改善了小鼠的糖耐量和全身胰岛素的敏感性[31]。这些研究结果证明脂肪组织在血糖稳态中能够起到关键的调控作用,而这一作用依赖于GluT4的功能。T2DM患者持续的高胰岛素导致脂肪组织GluT4表达下调,并抑制GluT4囊泡对胰岛素的响应[32],这可能造成T2DM患者进一步的胰岛素抵抗。
4 GluT在骨骼肌中的作用
骨骼肌是机体最大的能量消耗场所和胰岛素敏感组织,其能量代谢方式复杂,并受到精细的调节。休息状态下,骨骼肌的主要能量来源是FFA;餐后或运动后,骨骼肌可以摄取葡萄糖并合成肌糖原储存;运动或饥饿时,肌糖原反应性分解供能。餐后胰岛素作用下,由骨骼肌处理的葡萄糖约占70%~85%[33]。骨骼肌也是机体最大的糖原储存器官,但由于缺乏葡萄糖-6-磷酸酶,肌糖原只能分解供能,无法再产生葡萄糖进入血液。
葡萄糖转运是骨骼肌吸收利用葡萄糖的限速步骤。骨骼肌细胞表面的GluT也是胰岛素敏感的GluT4,在受到胰岛素刺激或肌肉收缩刺激时可以迅速激活,将大量葡萄糖转运至细胞内。骨骼肌过表达GluT4的小鼠具有更强的血糖处理能力[34]。骨骼肌特异性敲除GluT4则阻断了小鼠骨骼肌的葡萄糖摄取,导致糖耐量受损和IR[35,36]。骨骼肌特异性敲除胰岛素受体的小鼠丧失骨骼肌胰岛素响应,但其脂肪胰岛素敏感性代偿性提高、脂沉积增加、血浆甘油三酯和FFA水平升高,小鼠具有正常的葡萄糖耐量[37,38]。这说明骨骼肌中还存在不依赖于胰岛素的葡萄糖摄取信号通路。进一步研究发现,肌肉收缩刺激可以通过Ca2+浓度或腺苷酸活化蛋白激酶(AMP-activated protein kinase, AMPK)通路激活GluT4向膜的转位[27]。
肌糖原缺乏是T2DM患者骨骼肌的特征之一,糖原合成的减少是由葡萄糖转运效率下降和GK活性的改变造成的。T2DM患者的骨骼肌GluT4总蛋白水平和运动介导的葡萄糖摄取能力正常,而胰岛素介导的葡萄糖摄取能力降低,这说明可能是其胰岛素信号转导系统的缺陷导致了GluT4转位受阻,从而影响了骨骼肌葡萄糖摄取及糖原合成。研究表明,适度锻炼可以提高骨骼肌的胰岛素敏感性[39],从而缓解部分早期T2DM症状。
5 GluT在肠道中的作用
消化道中的葡萄糖主要依靠钠葡萄糖同向转运体(sodium glucose cotransporter,SGLT)的主动运输进入小肠黏膜上皮细胞,经过细胞内运输后,再借助于基底膜上GluT2的协助扩散作用被动地运送到全身[40]。当小肠中的葡萄糖浓度超过SGLT的最大转运能力后,小肠黏膜上皮细胞的GluT2也可以分布至顶膜,参与葡萄糖的吸收[41];肠腔内升高的葡萄糖通过甜味受体亚基(T1R3)、α-味转导素(α-gustducin)以及AMPK信号通路调节GluT2的表达和囊泡运输[42]。
小肠GluT2的功能除介导葡萄糖的吸收和扩散外,还在调节小肠微绒毛长度、L细胞功能、肠道通透性和炎症反应中发挥作用[43]。首先,GluT2小肠特异性敲除减弱了近端小肠对葡萄糖的吸收、延迟葡萄糖向肠道周围组织的分布、减少微绒毛的长度,从而模拟营养吸收不良的状态。其次,近端葡萄糖吸收的减少会增加葡萄糖向远端肠道的输送,从而为远端肠道微生物群提供新的可发酵能量来源;这些细菌可通过产生丁酸盐减少促炎细胞因子的表达,降低肠道通透性,对维持肠道稳态发挥积极作用。另外,GluT2的缺失虽然使肠道L细胞密度降低,但同时加强了L细胞胰高血糖素样肽-1(GLP-1)的分泌,因而不影响血液中GLP-1的含量。因此,通过药物特异性地阻断小肠GluT2活性或可为防止体重增加和代谢紊乱提供新思路。
6 GluT在神经系统与大脑中的作用
大脑不储存糖原和甘油三酯等能量,但需要持续的能量供应。大脑能够直接利用葡萄糖,但不能直接利用FFA。早期饥饿状态下,肝糖原分解和糖异生产生葡萄糖入血,并优先保证大脑的利用,此时葡萄糖仍是中枢神经系统的唯一燃料;其他组织葡萄糖氧化利用减少,转而利用FFA供能。长期饥饿状态下,大脑则可以利用由FFA转化而来的酮体提供能量。
动物大脑的葡萄糖摄取主要依靠GluT1和GluT3。GluT1在构成血脑屏障的微血管内皮细胞和除神经元外的脑实质表达[11],Km值约为6.9 mmol/L,与基础血糖相近。GluT1负责控制葡萄糖由血液循环稳定地通过血脑屏障进入脑脊液,以保证大脑充足、稳定的葡萄糖流量。GluT1缺陷综合症患者由于大脑的葡萄糖供应不足,会出现包括小儿耐药性癫痫、发育迟缓、痉挛、共济失调、复杂运动障碍等[44]。GluT3主要在神经元表达,相较于其他3个Ⅰ组GluT成员具有最高的底物亲和力(Km≈1.4 mmol/L)和较大的转运效率[11]。脑脊液的葡萄糖浓度平均只有1~2 mmol/L,远低于循环系统中的平均葡萄糖浓度,因此,GluT1和GluT3的精妙组合既能消除循环系统血糖瞬时升高对大脑的冲击,又可以确保神经元对于葡萄糖的优先使用权,且始终有充足的能量供应。
GluT除具有维持大脑和神经系统能量供应的功能之外,还在神经系统的葡萄糖感受中起到重要的作用。GluT2在下丘脑、脑干、脑室内神经元、星形胶质细胞以及肝门静脉内葡萄糖感受器中都有表达。GluT2依赖的葡萄糖敏感神经元可以通过迷走神经和交感神经控制动物的采食、体温调节以及胰岛的增殖与功能。首先,中枢葡萄糖感受器的作用依赖GluT2的表达[45, 46]。下丘脑弓状核能感知葡萄糖浓度的升高,并通过分泌神经肽Y(NPY)调节食欲、产热和胰岛素释放。进一步研究发现,虽然NPY神经元不表达GluT2,但与表达GluT2的神经元存在突触连接。中枢中GluT2依赖的葡萄糖感受器通过黑皮质素途径影响NPY神经元对于瘦素的敏感性,调控采食和产热[47]。其次,GluT2也参与植物性神经对于胰岛功能的调控。神经特异性GluT2敲除小鼠(NG2KO)的副交感神经放电频率下降,且腹腔葡萄糖注射无法引起副交感神经的激活或交感神经的抑制。NG2KO小鼠哺乳期的β细胞增殖相对于对照组减少了一半,并造成成年后的β细胞数减少了30%,说明副交感神经在促进细胞增殖及维持成年β细胞数量方面具有重要的作用,而GluT2的功能在其中是不可或缺的[48, 49]。最后,肝门静脉存在丰富的迷走神经传入支,当肝门静脉葡萄糖浓度升高时,其中的葡萄糖感受器被激活,引起第一相胰岛素分泌,抑制胰高血糖素分泌,并引起食欲抑制和骨骼肌与脂肪的糖摄取增强[50]。GluT2敲除鼠失去血糖对胰高血糖素分泌的调控,通过神经节阻滞可以恢复胰高血糖素的水平[14],表明门静脉葡萄糖感受器的功能依赖于GluT2的表达。
7 展望
综上所述,Ⅰ组GluT成员对于机体血糖的稳态调节至关重要,它们的异常表达会导致器官或组织的功能紊乱,与T2DM等代谢疾病甚至肿瘤的发生密切相关。当下,科学家们正致力于分析疾病发生过程中GluT表达模式的变化,将其运用于疾病诊断,并开发相应的治疗方法;在针对GluT开发药物时,不能忽视GluT在各类组织中所发挥的独特作用,以及GluT之间可能存在的相互代偿表达的关系。另外,目前仍缺少理想的大动物模型用于T2DM等慢性代谢疾病的临床研究;GluT作为糖代谢过程中的关键蛋白,可能成为构建代谢疾病大动物模型时的重要基因修饰位点。从调控血糖稳态关键组织的功能出发解析GluT在其中发挥的作用,有助于我们更直观地了解GluT家族各成员的特性与其分布组织器官的关系,以及相关代谢通路在血糖稳态调控中的作用。GluT及其相关生物学过程作为已知或潜在的药物靶点,为T2DM等代谢疾病的诊断和治疗提供了新的思路,值得进行更深入的研究和探讨。
