高温胁迫对樟树光合性能的影响
2019-01-24田正凤应彬彬马元丹左照江
王 彬,田正凤,应彬彬,马元丹,左照江
(浙江农林大学 林业与生物技术学院,浙江 杭州 311300)
在全球气候变暖的大背景下,由温室效应不断加剧而导致的高温天气日趋严重,已成为全世界广为关注的环境问题之一。高温不仅会危害人体健康,也会对农林业生产造成严重影响[1-2]。在高温胁迫下,植物种子萌发受抑[3]、 光合能力降低[4-5]、 活性氧(reactive oxygen species,ROS)大量产生并造成次生氧化胁迫[6]。这不仅会影响植物生长发育,甚至会引发细胞程序性死亡[6-7]。光合作用是植物最主要的物质与能量代谢过程,也是对环境因子变化最为敏感的生理过程[8-9]。高温胁迫可破坏光系统Ⅱ(PSⅡ)结构,抑制电子供体侧水的光解,并降低气孔导度、反应中心光化学效率、二氧化碳同化能力和水分利用效率等[4,10-12]。在这些影响中,往往几种共同作用以降低植物的光合性能。樟树Cinnamomum camphora主要生长在热带和亚热带地区,是优良的园林绿化树种,被广泛作为庭荫树、行道树、防护林以及风景林使用,具有巨大的经济与生态价值。目前,对樟树研究主要集中于组织培养和无性繁殖等育苗技术[13-14],以及低温[15]、 干旱[16-17]和盐碱胁迫[18-19]等对生理特性的影响以评估其向北方和盐碱滩涂引种的适应性。随着高温天气的加剧,其生长过程中不可避免的会经历高温胁迫,以其主要生长区域之一浙江省为例,2013-2017年间夏季高温可达40℃,甚至某些区域会高达42℃[20]。然而高温对樟树生长与光合作用具有何种影响,尚未见相关研究报道。因此,本研究从叶绿素荧光诱导动力学、气体交换速率和水分利用效率等方面研究高温胁迫对樟树光合性能的影响,以期揭示高温环境对樟树的危害。
1 材料与方法
1.1 实验材料与处理
在3月选取生长一致的3年生樟树幼苗(高度约90 cm)移栽至花盆(高30 cm,直径30 cm)中,并置于室外缓苗备用。在5月选取生长状况一致的幼苗进行高温胁迫处理。将樟树幼苗放入光照培养箱中(光16 h/暗8 h),分别进行25,35和45℃(光照时的温度)温度处理,黑暗时温度分别为20,25和35℃。35和45℃高温处理前分别经25和35℃适应2 d后进行,温度升高时升速为3℃·h-1。以25℃处理作为对照。每个温度处理幼苗4株,视为重复1次·株-1。高温胁迫2 d后,测定幼苗的光合性能。
1.2 叶绿素荧光诱导动力学曲线测定
对樟树叶片暗适应15 min后,采用Yaxin-1161叶绿素快速荧光仪测定其叶绿素荧光诱导动力学曲线[21],并根据CHRISTEN等[22]的方法计算叶绿素荧光动力学参数:单位反应中心吸收的光能总量(ABS/RC),单位反应中心捕获的光能总量(TRO/RC),单位反应中心内电子传递的量子产额(ETO/RC),单位反应中心热耗散的能量(DIO/RC),光合性能指数(PIABS)和单位吸收面积上天线色素能量吸收的驱动力(DFABS)。
1.3 气体交换速率测定
樟树叶片的气体交换速率采用LI-6400便携式光合仪进行测量,光强设定为2 000,1 500,1 200,1 000,800,600,400, 300, 200,100,50, 20,10 和 0 μmol·m-2·s-1,二氧化碳摩尔分数为 400 μmol·mol-1, 叶室温度为 30 ℃。 对光合速率(Pn), 气孔导度(Gs), 胞间二氧化碳摩尔分数(Ci)和蒸腾速率(Tr)的光响应变化进行分析,并参考YE等[23]直角双曲线修正模型计算光补偿点、光饱和点和最大光合速率。
1.4 水分利用效率计算
水分利用效率是指植物或叶片每蒸腾一定量的水分所同化的二氧化碳的量,即Pn与Tr的比值,以mmol·mol-1(二氧化碳/水)表示。 在 25和 35℃时, 樟树处于最大光合速率时的光强接近于 1 200 μmol·m-2·s-1, 45 ℃时接近 1 000 μmol·m-2·s-1, 分别采用此时的光合速率和蒸腾速率计算相应的水分利用效率。
1.5 数据处理
实验数据采用SPSS 13.0进行分析,Origin 8.0进行作图。
2 结果与分析
2.1 高温胁迫对樟树叶绿素荧光诱导动力学的影响
在35和45℃高温胁迫下,樟树叶绿素荧光诱导动力学曲线O到P点的荧光强度均明显降低,其中在45℃高温胁迫后降至最低(图1)。
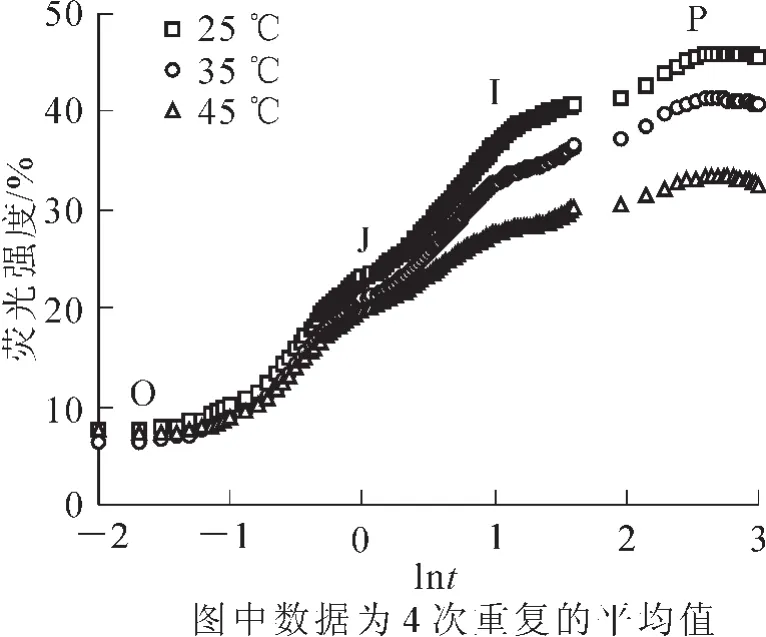
图1 高温对樟树叶绿素荧光诱导动力学曲线的影响Figure 1 Effects of high temperature on chlorophyll fluorescence transientsin Cinnamomum camphora
樟树经35和45℃高温胁迫处理后,其ABS/RC分别降低了4.9%和21.7%,其中在45℃高温胁迫时差异达到极显著(P<0.01)水平。与ABS/RC相似,35和45℃高温胁迫均明显降低TRO/RC,ETO/RC,PIABS和DFABS,并且在45℃高温胁迫时均达到极显著(P<0.01)水平。对于DIO/RC而言,35和45℃高温胁迫均使其明显增加,与对照相比分别增加了13.5%(P<0.05)和 78.4%(P<0.01)(表 1)。
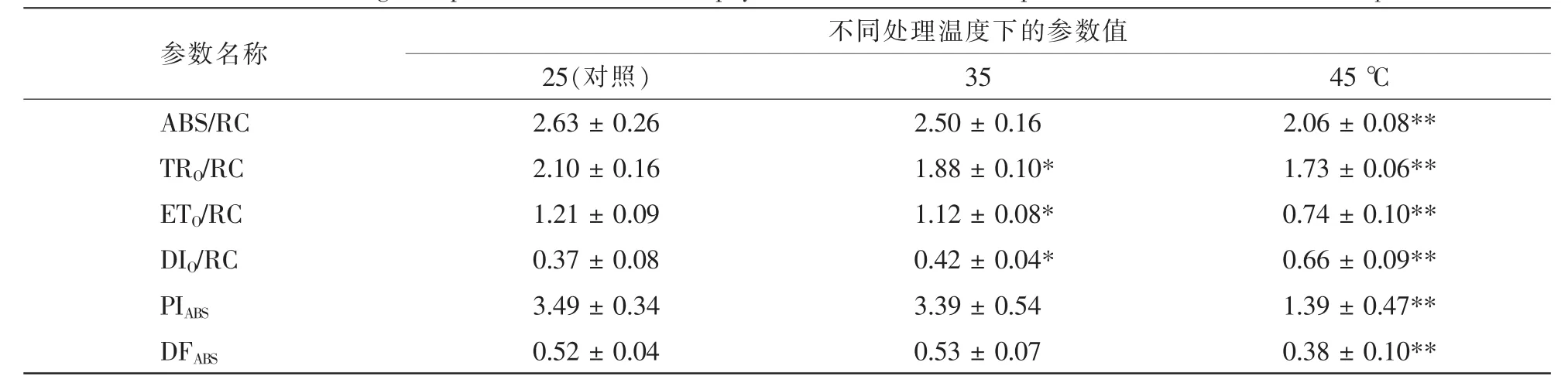
表1 高温胁迫对樟树叶绿素荧光诱导动力学参数的影响Table 1 Effects of high temperature stress on chlorophyll fluorescence transient parameters in Cinnamomum camphora
2.2 高温胁迫对樟树光合作用的影响
随着光照强度增加,樟树光合速率呈现先增加后趋于稳定的明显的光响应曲线特征。在35和45℃高温胁迫后,樟树的光响应曲线明显降低,其中在45℃时降至最低,这表明高温明显抑制了樟树光合速率(图2A),其中最大光合速率分别降低了42.3%(P<0.05)和67.6%(P<0.05)。 同时, 光补偿点分别增加了20.3%(P<0.01)和1.6倍(P<0.01),而光饱和点则分别降低了6.7%和11.9%(P<0.05)(表2)。
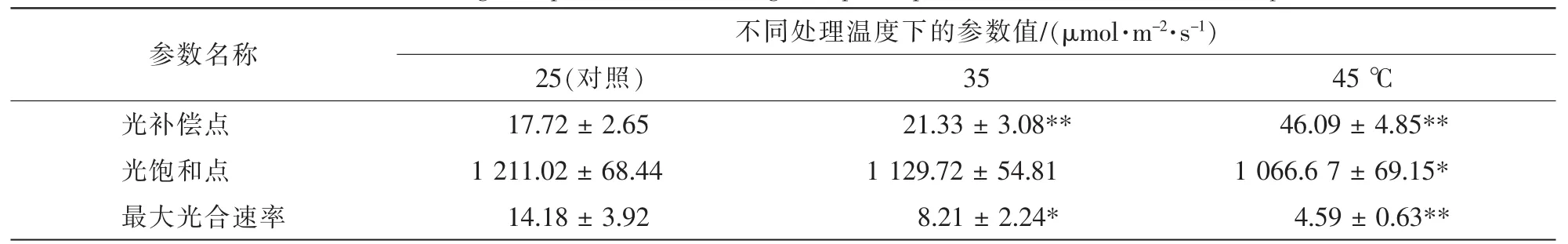
表2 高温胁迫对樟树光响应参数的影响Table 2 Effects of high temperature stress on light response parameters in Cinnamomum camphora
气孔导度对光强的响应与光合速率相似,亦随光强增加而呈现先增加后趋于稳定的变化趋势,同时随高温胁迫增强而逐渐降低(图2B)。对于胞间二氧化碳摩尔分数而言,其随光强增加而逐渐降低。在高温胁迫后, 胞间二氧化碳摩尔分数升高, 其中在光强≥1 000 μmol·m-2·s-1后 45 ℃时最高(图 2C)。

图2 高温胁迫对樟树光合作用的影响Figure 2 Effects of high temperature stress on photosynthesis in Cinnamomum camphora
2.3 高温胁迫对樟树蒸腾速率和水分利用效率的影响
随着光照强度增加,樟树在25,35和45℃的蒸腾速率均呈现逐渐增加趋势。与25℃相比,樟树在35和45℃高温胁迫后,其蒸腾速率明显降低,其中在45℃高温胁迫时降至最低(图3A)。采用樟树处于最大光合速率时的光合速率和蒸腾速率计算水分利用效率,其在25℃时为5.0 μmol·mmol-1,在35和45℃高温胁迫后分别降低了16.0%和44.6%,并且差异均达到显著(P<0.05)和极显著(P<0.01)水平(图3B)。
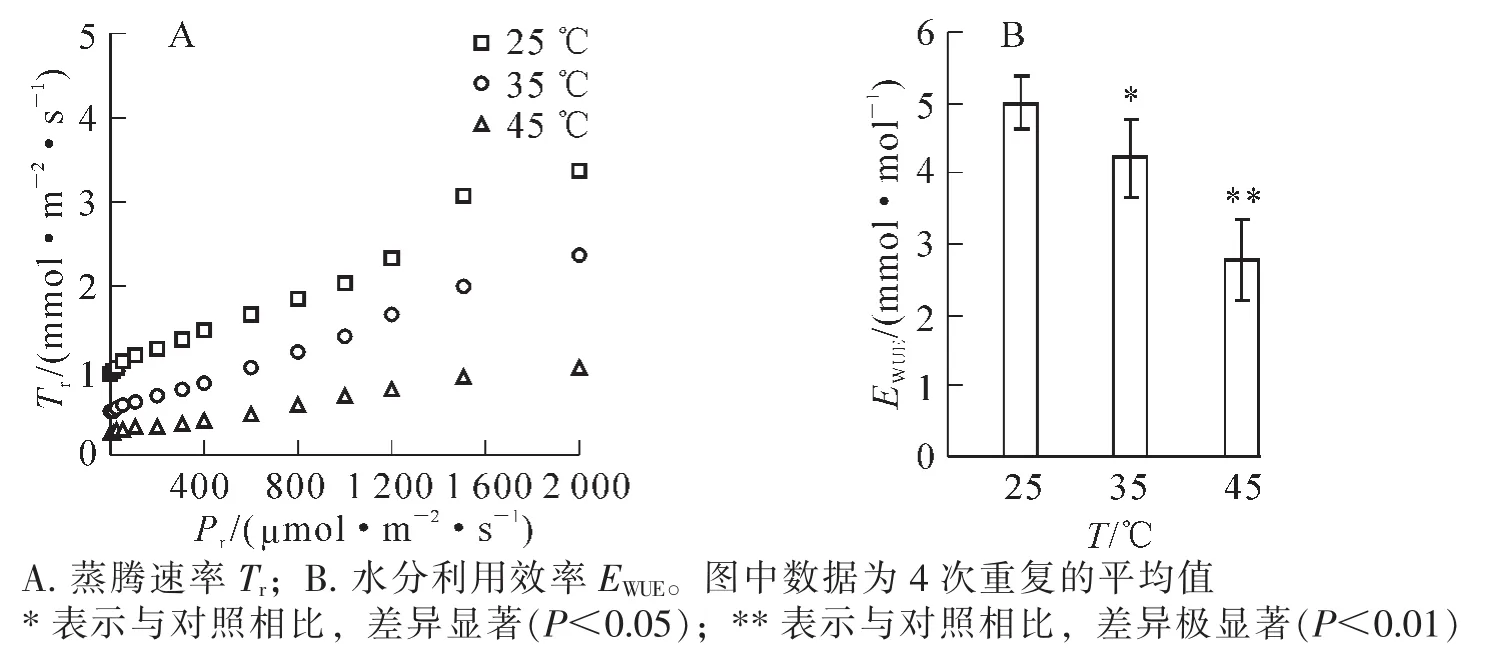
图3 高温胁迫对樟树蒸腾速率和水分利用效率的影响Figure 3 Effects of high temperature stress on transpiration rate and water use efficiency in Cinnamomum camphora
3 讨论
叶绿素荧光是研究植物光合作用有效的非损伤探针,其诱导动力学曲线可反映PSⅡ反应中心原初光化学反应。在O—P点的荧光动态变化过程中,O点表示PSⅡ反应中心可最大限度的接受光量子;O—J表示PSⅡ反应中心受光激发后将电子传给PSⅡ原初醌受体(QA)并还原为;J—I—P表示质体醌(PQ)库的还原状态[24]。在高温胁迫下,樟树O—P点的荧光强度明显降低,这与高温胁迫对苹果Malus domestica[25]和黄瓜Cucumis sativus[26]叶片叶绿素荧光诱导动力学曲线的影响相一致。在此过程中,樟树O—J点荧光强度降低,表明其积累导致了PSⅡ受体侧电子传递受阻[21,27];J—P降低则表明PSⅡ供体侧电子传递受阻, 并引发 P680+积累[28-29]。
在光反应过程中,光能的吸收、分配与利用可通过叶绿素荧光诱导动力学参数进行反应[30]。在高温胁迫下,辣椒Capsicum annum叶片反应中心单位面积上吸收、捕获和传递的光能均明显降低[31]。在本研究中,樟树亦有相似结果,ABS/RC,TRO/RC和ETO/RC在高温胁迫下均明显低于对照(表1)。在高温胁迫时,樟树DIO/RC增加,这表明其吸收的光能通过叶黄素循环进行热耗散增加[33]。PIABS由光能吸收、激发能捕获和激发能到电能的传导3方面构成[32],在高温胁迫下樟树PIABS降低表明光反应过程受到明显抑制。
在高温胁迫后, 黄瓜[34]、 芍药 Paeonia lactiflora[35]和油菜 Brassica napus[36]的光合速率、气孔导度和蒸腾速率以及芍药水分利用效率均明显降低,同时黄瓜[34]和西洋杜鹃Rhododendron hybridum[37]胞间二氧化碳摩尔分数明显升高,本研究结果与之相似。樟树在高温胁迫后,气孔导度降低限制了水分蒸发从而导致蒸腾速率降低,同时光合速率降低量明显高于蒸腾速率降低量,进而导致水分利用效率明显降低。植物光合速率降低的原因可分为气孔限制与非气孔限制[38],如果胁迫降低气孔导度而叶肉细胞仍可活跃的进行光合作用,其胞间二氧化碳摩尔分数显著降低,这就是典型的气孔限制;反之则为非气孔限制。在高温胁迫下,虽然樟树气孔导度明显降低,但是胞间二氧化碳摩尔分数却明显升高,这表明高温导致樟树光合速率降低不是由气孔限制原因造成的。
由此可见,高温胁迫可通过降低樟树PSⅡ的光能吸收、量子产量和电子传递,并促进吸收的光能进行热耗散,从而降低PSⅡ效率,进而导致同化力(三磷酸腺苷ATP和还原型烟酰胺腺嘌呤二核苷酸磷酸NADPH)减少以影响二氧化碳同化并降低光合速率。
