干旱胁迫及复水对入侵植物节节麦幼苗生长及生理特性的影响
2019-01-23王宁袁美丽陈浩李真真张铭鑫
王宁,袁美丽,陈浩,李真真,张铭鑫
(1.河南科技大学林学院,河南 洛阳 471023;2.洛阳市隋唐城遗址植物园,河南 洛阳471002)
节节麦(Aegilopstauschii),禾本科山羊草属植物,最早起源于西亚及东欧等地,现已成为世界恶性杂草之一[1]。自1955年在我国河南省新乡地区首次被发现,至今已广泛入侵至渭河流域陕西、甘肃,黄河流域河北、河南、山东以及内蒙古等地[2]。因节节麦具有生长适应能力强,耐瘠薄、分蘖系数高、出苗时间长等特点,又与小麦(Triticumaestivum)籽粒大小、形状较为相似,且无有效防除的除草剂,导致其成为麦田中最难防除的恶性杂草之一。据报道,仅2003年河北南部地区由于节节麦危害导致的小麦损失就达10%~25%[3]。因严重威胁粮食的生产与安全,早在2007年节节麦已被列入我国首次发布的《中华人民共和国进境植物检疫性有害生物名录》,如今许多国家及地区也已将其列入粮食贸易中的重点检疫对象[4]。
为提高自身的竞争及入侵能力,外来植物常表现出较高的形态及生理可塑性,以适应不同的生境条件[5]。如入侵植物飞机草(Eupatoriumodoratum)依据超氧化物岐化酶(superoxide dismutase, SOD)、抗坏血酸过氧化物酶(ascorbate peroxidase, APX)等保护酶活性的持续增加[6],以及较高的根冠比和水分利用效率[7],从而具有一定耐旱性,这也促进了其向干旱及半干旱地区的入侵。但宋莉英等[8]研究发现,三裂叶蟛蜞菊(Wedeliatrilobata)的光合作用对干旱胁迫较敏感,认为水分可能是影响其生长及扩散的重要限制因子。土壤水分是影响植物生长发育及分布范围的重要限制性因子[9]。因此,开展外来植物的耐旱性研究,对预测其分布范围及进一步扩散的防控具有十分重要的意义。众多研究认为,外来植物的形态结构、生物量积累与分配以及生理代谢等均会受到干旱胁迫的影响[6-7,10-12]。而植物的耐旱性不仅表现在干旱的胁迫过程中,更重要的是复水后的恢复过程,因此,能否在复水过程中迅速弥补之前遭受的损伤是植物对干旱胁迫适应的重要表现[13-14]。本试验通过盆栽试验,设置干旱-复水条件,依据干旱过程中形态、生理参数,以及复水过程中生理参数的变化,探讨节节麦对土壤干旱胁迫的适应机制,以期为节节麦入侵机制研究提供理论依据。
1 材料与方法
1.1 材料
2017年5月,节节麦种子采自河南省新乡市农业科学院试验田。游标卡尺测得种子平均纵径为5.98 mm,横径为2.54 mm,万分之一天平测千粒重为12.32 g。试验在河南科技大学开元校区林学院试验地内进行。研究区域位于洛阳市洛龙区,地理位置为34°59′ N、112°16′ E。试验所用土壤为褐土,来源于洛阳市洛龙区周边农田,基础理化性状为:pH值7.4,全氮含量0.85 g·kg-1,有效磷11.4 mg·kg-1,速效钾167.5 mg·kg-1。
1.2 试验设计
2018年2月,节节麦种子进行催芽处理,待幼苗长至“两叶一心”时进行移栽上盆(盆口Φ 30 cm,高25 cm),6株/盆。盆土提前经105 ℃烘箱烘干至恒重,干土5 kg/盆。土壤水分设置包括:正常水平NL(normal level,田间最大持水量的75%~80%)、轻度干旱LD(light drought,田间最大持水量的60%~70%)、中度干旱MD(moderate drought,田间最大持水量的55%~60%)、重度干旱HD(heavy drought,田间最大持水量的45%~50%)4个水分梯度[15-16],每个处理8盆,5次重复。缓苗1周后正常培养,待幼苗长至“四叶一心”时开始进行土壤控水处理。胁迫期间,采用称重法[17]控制土壤含水量,称重1次/2 d,当土壤含水量低于设置范围时,采用缓慢浇水法补充水分。分别在胁迫处理的0(CK)、7、14、21、28 d进行采样(从上至下第3~4片叶),第28天取样后复水至CK水平,并在复水第7天(即处理第35天)再次采样,进行生理指标的测定。其中,在干旱胁迫的第28天,每处理随机选出5株幼苗进行生物量的测定,利用扫描仪、Photoshop软件进行叶面积的测定[18],之后将植株的叶、茎及根分开并装纸袋,于80 ℃烘箱中烘干至恒重。
1.3 测定项目及方法
生理参数包括:采用称重法测定叶片相对含水量(relative water content,RWC)[19];采用95%乙醇法测定叶绿素含量,采用酸性茚三酮法测定脯氨酸含量,采用氮蓝四唑光还原法测SOD活性,采用硫代巴比妥酸法测定丙二醛(malondialdehyde, MDA)含量[20];采用电导法测定相对电导率(relative conductivity,REC)[21]。形态参数包括:叶生物量比(叶干重/植株总干重,leaf mass ratio,LMR);根生物量比(根干重/植株总干重,root mass ratio,RMR);比叶面积(总叶面积/总叶重,specific leaf area,SLA);根冠比(根干重/植株地上部分干重,root-shoot ratio,R/T)[22]。此外,可塑性指数(plasticity index,PI)=[(不同处理下某参数的最大值-最小值)/最大值][23]。
1.4 数据分析
采用SPSS 18.0进行数据分析,单因素方差分析和Duncan多重比较分析同一胁迫水平不同胁迫时间各个参数的差异显著性。图中数据为平均值±标准差。
2 结果与分析
2.1 干旱胁迫及复水对节节麦生理参数的影响
2.1.1干旱胁迫及复水对节节麦相对含水量的影响 随胁迫时间的延长,NL的RWC基本保持稳定,而各干旱处理的RWC均呈下降变化趋势(图1)。其中,LD的RWC至胁迫28 d时与CK差异未达到显著水平,而MD、HD的RWC分别在胁迫的21及14 d时与CK差异达到显著水平(P<0.05)。复水后,各干旱处理的RWC均呈升高变化,且与CK差异均不显著。
2.1.2干旱胁迫及复水对节节麦叶绿素含量的影响 如图2所示,随胁迫时间的延长,各干旱处理的叶绿素含量整体呈先升后降的变化趋势。其中LD、MD的叶绿素含量峰值均出现在胁迫的21 d时,HD的叶绿素含量峰值出现在胁迫的7 d时,但与CK差异均不显著;之后,叶绿素含量呈下降变化,至胁迫28 d时,仅HD的叶绿素含量与CK差异达到显著水平(P<0.05)。复水后,各处理的叶绿素含量均呈增加变化,但与CK差异均未达到显著水平。
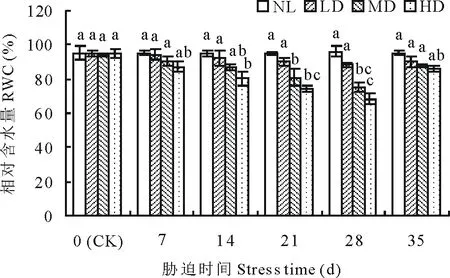
图1 干旱胁迫及复水对节节麦叶片相对含水量的影响Fig.1 Effects of drought stress and rewatering on the leaf relative water content (RWC) of A. tauschii
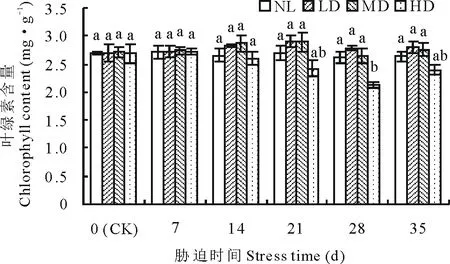
图2 干旱胁迫及复水对节节麦叶绿素含量的影响Fig.2 Effects of drought stress and rewatering on the leaf chlorophyll content of A. tauschii
NL, LD, MD, HD分别表示正常水平、轻度干旱、中度干旱、重度干旱。不同小写字母表示差异显著(P<0.05),下同。NL, LD, MD, HD indicate normal level, light drought, moderate drought and heavy drought. The different lowercase letters mean significant difference at 0.05 level, the same below.
2.1.3干旱胁迫及复水对节节麦相对电导率的影响 随胁迫时间的延长,各干旱处理的相对电导率均呈持续增加的变化趋势(图3)。其中,胁迫至28 d时LD的REC较CK增加了18.91%,但差异未达到显著水平;MD、HD的REC分别在胁迫28和21 d时与CK差异达到显著水平(P<0.05)。复水后,REC均明显下降,其中,MD、HD的REC仍高于CK,但差异均不显著(P>0.05)。
2.1.4干旱胁迫及复水对节节麦丙二醛含量的影响 各干旱胁迫处理中,MDA含量均呈缓慢增加变化趋势(图4)。其中,LD的MDA含量至胁迫的28 d时较CK增加了11.97%,但未达到差异显著水平;MD、HD的MDA含量分别在胁迫的21和14 d时与CK差异达到显著水平(P<0.05)。复水后,各处理的MDA含量均明显下降,其中,MD、HD的MDA含量明显高于CK,但差异均未达到显著水平。

图3 干旱胁迫及复水对节节麦相对电导率的影响Fig.3 Effects of drought stress and rewatering on the relative conductivity of A. tauschii
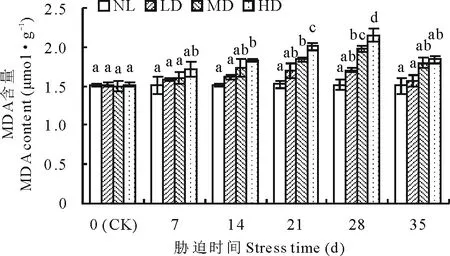
图4 干旱胁迫及复水对节节麦MDA含量的影响Fig.4 Effects of drought stress and rewatering on MDA content of A. tauschii
2.1.5干旱胁迫及复水对节节麦脯氨酸含量的影响 各干旱处理中,脯氨酸含量均呈持续增加的变化趋势(图5)。其中,LD、MD的脯氨酸含量在胁迫的14 d时与CK差异均达到显著水平(P<0.05),而HD的脯氨酸含量则是在胁迫的7 d时,与CK差异显著(P<0.05)。复水后,脯氨酸含量均明显下降,但仍均高于CK,其中LD、MD的脯氨酸含量与CK差异均不显著,而HD的脯氨酸含量则依然显著高于CK(P<0.05)。
2.1.6干旱胁迫及复水对节节麦超氧化物歧化酶活性的影响 随胁迫时间的延长,LD、MD的SOD活性呈持续增加变化趋势,而HD的SOD活性则呈先升后降的变化趋势(图6)。 其中,LD、MD的SOD活性分别在胁迫的21及14 d与CK差异达到显著水平(P<0.05),胁迫至28 d时,分别较CK增加了44.33%、59.01%;而HD的SOD活性在胁迫的7 d时与CK差异达到显著水平(P<0.05),并在胁迫的21 d时达到峰值,较CK增加了63.98%,之后SOD活性略有下降。复水后,LD、MD的SOD活性迅速下降,但与CK差异均不显著,而HD的SOD活性则略有升高,且与CK差异显著(P<0.05)。
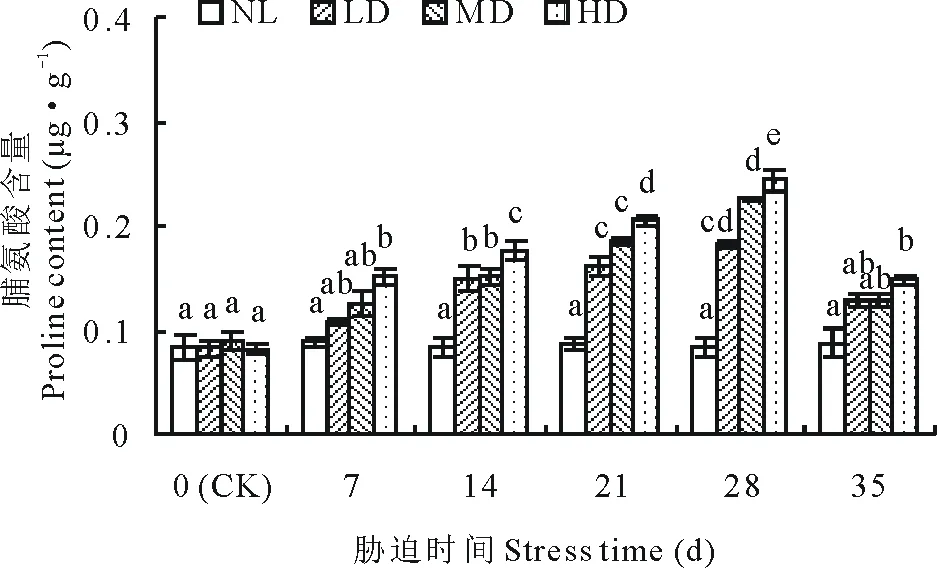
图5 干旱胁迫及复水对节节麦脯氨酸的影响Fig.5 Effects of drought stress and rewatering on proline content of A. tauschii
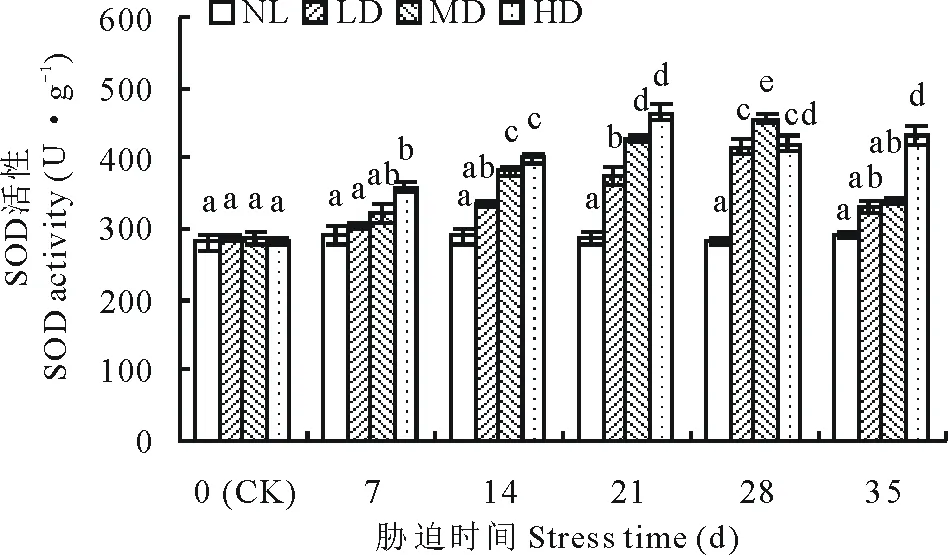
图6 干旱胁迫及复水对节节麦SOD活性的影响Fig.6 Effects of drought stress and rewatering on SOD activity of A. tauschii
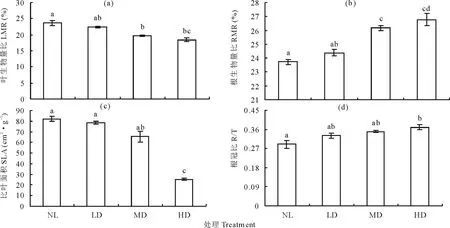
图7 干旱胁迫对节节麦形态指标的影响Fig.7 Effects of drought stress on morphological index of A. tauschii
2.2 干旱胁迫对节节麦形态特征的影响
随土壤含水量的降低,LMR、SLA逐渐下降,而RMR、R/T则呈上升变化趋势。其中,当土壤含水量下降至MD时,LMR、RMR均与NL差异显著(P<0.05);土壤含水量降至HD时,SLA、R/T均与NL差异显著(P<0.05)。
2.3 干旱胁迫对节节麦形态及生理参数可塑性指数的影响
从表1可知,生理参数中,以Pro的PI值最大,叶绿素含量的PI最小;形态参数中,以SLA的PI值最大,RMR的PI最小。且形态参数PI的均值为0.31,明显小于生理参数的0.39。
3 讨论
3.1 干旱胁迫及复水对节节麦生理代谢的影响
RWC是植物体内水分状况的重要反映指标,一定程度上能体现出植物对干旱胁迫的适应能力,因此常被用于植物耐旱性的鉴定[24]。干旱胁迫中,RWC降速慢且降幅小,表明该植物的持水保水能力强,即具有一定的耐旱性[25]。本研究中,干旱胁迫中RWC的下降与土壤含水量及可利用水的减少有关,造成节节麦根系吸水困难,再有叶片的蒸腾丧失,从而导致了RWC的下降,复水后各处理的RWC迅速恢复至正常水平,可见节节麦叶片具有较强的保水持水能力,这也为其能够在干旱胁迫下维持正常的生理代谢奠定了基础,这也与宋家壮等[24]、郭颖等[26]对强耐旱植物的研究结论一致。
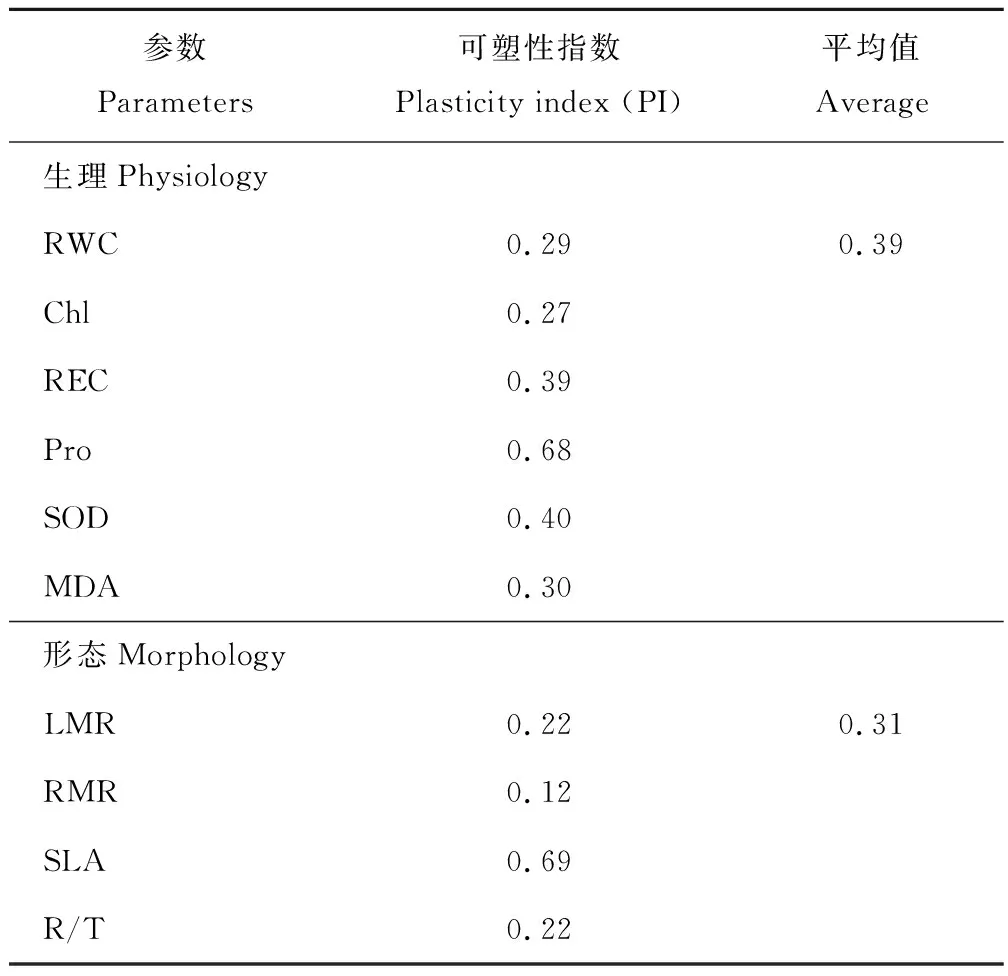
表1 干旱胁迫对节节麦形态及生理参数可塑性指数的影响Table 1 Plasticity index for each of the morphological characteristics and physiology of A. tauschii under drought stress
注:LMR, RMR, SLA, R/T分别表示叶生物量比、根生物量比、比叶面积、根冠比;RWC, Chl, REC, Pro, SOD, MDA分别表示相对含水量、叶绿素含量、相对电导率、脯氨酸含量、超氧化物歧化酶、丙二醛。
Note: LMR, RMR, LAR, R/T indicate leaf mass ratio, root mass ratio, specific leaf area and root-shoot ratio. RWC, Chl, REC, Pro, SOD, MDA indicate relative water content, chlorophyll, relative conductivity, proline, superoxide dismutase, malondialdehyde.
叶绿素含量的变化一定程度上可反映植物光合作用的变化规律,因此,可作为植物对水分胁迫敏感性的指示性指标[27]。多数研究认为,干旱胁迫会造成植物叶绿素含量的降低[28-30]。但宋家壮等[24]研究发现,干旱胁迫下虉草(Phalarisarundinacea)叶绿素含量呈增加的变化趋势,分析认为可能由于水分胁迫造成叶片含水量下降而产生的浓度效应所致,这也与张晓海等[31]、梁国玲等[32]的研究结论相符。本研究中,干旱胁迫下,节节麦叶绿素含量整体呈先升后降的变化趋势,前期的升高与宋家壮等[24]研究结论一致,而之后的下降可能与持续干旱胁迫造成叶绿素分解加速及合成减缓有关。胁迫至28 d时,仅有HD的叶绿素含量较CK减少显著(P<0.05),复水后各处理均又迅速恢复至正常水平,由此说明叶绿素的合成并未受到干旱胁迫的根本性破坏,节节麦能够通过自身调节进行恢复。
REC反映了植物细胞膜透性,MDA是膜脂过氧化作用的重要产物,因此,REC和MDA是植物质膜受伤害程度的重要反映指标[33-34]。本研究中,各干旱处理下,REC和MDA含量均呈持续增加的变化趋势,表明节节麦细胞膜受胁迫的伤害程度逐渐增大,这也与墨西哥羽毛草(Nassellatenuissima)[34]、白三叶(Trifoliumrepens)[35]及草地早熟禾(Poapratensis)等[36]的研究结论一致。复水后,REC和MDA含量均迅速下降,且与CK差异均不显著,由此可见,通过自身的调节机制,节节麦对干旱胁迫造成的膜系统损伤进行了积极地修复,这也可能是节节麦对干旱胁迫的重要适应机制。
渗透调节物质含量的增加是植物对逆境胁迫的重要生理反映,而游离脯氨酸是植物体内重要的渗透调节物质之一[37],许多研究表明,脯氨酸的积累与植物的耐旱性呈正相关[13,38-39]。本研究中,干旱胁迫造成脯氨酸含量持续增加,由此表明节节麦幼苗通过脯氨酸含量的增加来降低细胞渗透势,保障其体内正常的代谢活动,从而抵御干旱胁迫,复水后,胁迫被解除,LD、MD的脯氨酸含量恢复至正常水平,而HD的脯氨酸含量依然显著高于CK(P<0.05),这可能是干旱胁迫致使节节麦渗透调节能力增强所致。
SOD是植物体内重要的保护酶,其活性大小是植物抗旱性强弱的重要体现[6,33]。干旱胁迫过程中,LD、MD及HD前期,SOD活性均迅速增加,表明节节麦具有较高的清除O2-的能力,之后HD中的SOD活性略有下降,这可能与胁迫时间过长,超出了其忍受限度而造成SOD活性下降有关,这与入侵植物五爪金龙(Ipomoeacairica)[40]、牛膝菊(Galinsogaparviflora)[41]的研究结论一致。复水后,LD、MD中SOD活性基本恢复至正常水平,而HD中SOD活性依然显著高于CK(P<0.05),表明重度干旱胁迫及复水处理提高了节节麦对O2-的清除能力,这与张彦妮等[39]对黄连花(Lysimachiadavurica)的研究结论一致。
3.2 干旱胁迫对节节麦形态特征的影响
植物对环境变化的响应不仅体现在形态特征上,同时也体现在生物量分配的对策方面[42]。生物量分配格局的改变是植物应对环境变化的自我调整策略之一[43]。随土壤含水量的降低,节节麦幼苗的LMR呈下降趋势,而RMR、R/T呈增加变化趋势,这与入侵植物五爪金龙[44]、黄顶菊(Flaveriabidentis)[45]、互花米草(Spartinaalterniflora)[46]等的研究结论一致。表明节节麦通过增加根生物量的分配比及根冠比,拓展地下空间以获取生存所需水分,同时减少地上部分的生物量分配比,即尽量缩小地上空间以减少水分散失,达到规避干旱胁迫的效果,从而拥有了一定的耐旱能力。此外,土壤含水量的减少致使SLA的下降,表明节节麦还通过减少水分蒸发的途径适应干旱胁迫,这与王林龙等[47]的研究结论一致。
4 结论
节节麦幼苗通过保护酶活性及渗透调节物质含量的增加适应土壤干旱胁迫,从而表现出一定的耐旱性,复水后,脯氨酸、叶片相对含水量等多数指标均能恢复至正常水平,说明节节麦对干旱胁迫造成的损伤具有较高的修复能力。此外,节节麦幼苗通过降低地上部分及增加地下部分的生物量分配比,以及减少水分蒸发途径等适应土壤干旱环境。节节麦形态参数的可塑性指数均值明显小于生理参数,由此表明,节节麦主要通过生理代谢的调整适应短期干旱胁迫。
