延胡索光合与生长可塑性对光照的响应
2019-01-10盛海燕李伟成
盛海燕, 李伟成
延胡索光合与生长可塑性对光照的响应
盛海燕1, 李伟成2, 3, *
1. 杭州环境保护科学研究院, 杭州 310005 2. 国家林业局竹子研究开发中心/浙江省竹子高效加工重点实验室, 杭州 310012 3. 西南林业大学生命科学学院, 昆明 650224
林-药复合系统是经济收益显著的立体林农生态系统。以早春植物延胡索为研究对象, 分析其不同光环境下光合与生长的变化, 为其林下抚育、林木间伐和林药复合生态系统的构建提供依据。试验发现, 延胡索通过降低光饱和点、光补偿点和暗呼吸速率, 提高表观量子效率等方式以适应遮阴和弱光环境; 在高光照和中高光照条件下, 延胡索对两种光环境表现差异不显著, 在暗呼吸速率和最大净光合速率方面, 中高光照条件的均值甚至超过高光照条件; 弱光条件下的饱和光照强度、最大净光合速率和表观量子效率分别为高光照的62.67%、43.25%和58.42%。遮阴条件的净光合速率、气孔导度和蒸腾速率总体均低于高光照条件。遮阴后, 延胡索叶片的Chl a、Chl b及Chl a+Chl b皆随着有效光辐射的下降呈现先升后降的趋势, Chl a/Chl b值在中高光照和中低光照条件下显著降低, 表现出耐阴性能, 而弱光条件下Chl a/Chl b值显著下降的原因是弱光导致延胡索的光合产物供应不足, 妨碍了叶绿素的合成。弱光下延胡索叶生物量、根生物量分别仅为高光照条件下的34.78%和22.03%; 随着有效光辐射的下降, 比叶面积先升后降, 在弱光条件下延胡索优先于叶片的形态构成, 而不再倾向于扩大叶面积捕获更多的光量子。延胡索耐阴能力介于阳性植物和阴性植物之间, 弱光下的延胡索光合机构可能已向耐阴植物转变。因此, 延胡索具有一定的耐阴能力, 但并不适合长期的弱光条件。野外疏林下进行仿野生延胡索栽培时, 当乔冠层的郁闭度过高时, 可适当采取间伐修枝或者开辟林窗等措施改善林下光环境, 提升林下延胡索的光合作用能力和光斑利用效率, 合理配置其光生态位和时间生态位以达到预期的经济效益。
延胡索; 早春植物; 弱光; 林-药复合生态系统; 生理生态
1 前言
植物整个生命周期都对光环境产生不同程度的响应[1], 光合作用决定了植物生存与繁衍[2], 故如何保证植物高效利用光能具有重要意义。时间和空间的变化常使植物处于非理想光环境中, 因此, 森林生态系统冠层遮挡导致的弱光情况普遍存在于植物生长过程中。
发展林下复合生态系统已成为增加林地产出、提升林业效益、解决“三农”问题的重要途径之一。林-药复合生态系统是目前应用较多和经济收益最为显著的一种模式, 其主要运用生态群落学的生态位和空间结构原理在林下采用仿野化模式种植草本和灌木类药用植物的一种立体林农生态系统。林-药复合生态系统中, 林下弱光环境是控制草本和灌木植物光合作用的关键因子[3]。不同植物种类或者不同微生境, 对弱光的生理响应有很大差异[4]。如喜阴植物绞股蓝, 当林下光环境约为自然光照30%—40%时, 有利于其光合作用的进行[5]; 吴德成等[6]发现, 不同面积人工林窗内的光环境对林下人参()的栽培差异并不显著; 而不同乔木林冠和不同大小林窗的弱光环境对南方红豆杉(var.)苗木的影响不同[3]; 帚石楠()和欧洲蕨()在采伐后林窗中的表现截然不同, 帚石楠对光环境较为敏感, 欧洲蕨更有潜力占领林窗[7]; 走马胎()的净光合速率、光饱和点、光补偿点及光下呼吸速率随着林下透光度递减而减弱, 20.2%的弱光环境最适合其苗木的生长[8]。
延胡索()属早春植物, 其块根是传统大宗中药, 浙江省栽培历史悠久, 适宜其生长的野外落叶阔叶林林下环境因人类干扰而逐渐缩小。目前, 延胡索的药用成分、药理临床和大田栽培技术的研究日趋成熟。调查自然分布延胡索的群落发现, 具有一定有效光辐射的毛竹林、常绿落叶混交林及阔叶落叶混交林的林下有延胡索群落分布。因此, 本研究以浙江省磐安县延胡索种源为研究对象, 分析不同光环境下延胡索光合特性的变化, 特别是弱光条件下的光合特征及对不同光环境的适应性研究, 以期获得延胡索生长对光环境的要求和响应的差异, 为森林抚育间伐和延胡索林药复合生态系统的构建提供数据支撑。
2 研究地自然概况
试验地位于浙江省中部的金华市磐安县, 本区气候属亚热带季风区, 温暖湿润, 四季分明, 多年平均气温为13.9—17.4 ℃, 年积温4255—5534 ℃。雨量充沛, 多年平均降水量1409.8—1527.8 mm。无霜期较短, 200—243 d。海拔600 m以下的地带性土壤为红壤, 此外还有潮土和水稻土。境内地形条件复杂, 山峦起伏, 森林覆盖率高, 构成了气候的多层次和复杂性, 大面积分布类型多样的天然次生林。
中药材浙八味之一的延胡索, 喜温暖气候和向阳、湿润的环境, 浙江省中部地区是其主要产区, 由于经济结构的调整, 延胡索种植面积减少和规模减弱, 因此, 为保证中药材市场及中成药生产企业对延胡索生药的需求, 构建延胡索的林药复合生态系统意义重大。
3 试验设计和数据处理
3.1 试验设计
2016年9—10月采用控根容器播种, 高40 cm, 直径35—40 cm, 于丘陵熟地(前茬为浙贝母)挖表层20 cm砂质土壤混匀装盆, 土壤有机质19.2 g·kg–1, 速效磷33.7 mg·kg–1, 速效钾60.4 mg·kg–1, pH5.3。每盆控根容器种植播种3粒, 种茎大小一致(种茎质量1.15±0.22 g), 播种后于每盆表面施竹炭肥100 g(竹炭35 wt.%, N11.9 wt.%, P2O54.8 wt.%, K2O5.7 wt.%, pH8.13—8.50, 电导率5.47 mS·cm–1, 含水率30.5%)[9]。根据对浙江省中部与北部地区野生延胡索的光环境调查, 于2016年11月设置4种光环境, 即高光照(大田对照, 100%全光照)、中高光照(单层蓝色纱网遮阴, 82.3%全光照)、中低光照(双层蓝色纱网遮阴网, 37.9%全光照)和弱光(黑色遮阴网, 13.8%全光照)。出苗后留苗1株, 2017年5月采收, 种植-采收过程约为210 d左右, 期间常规管理。
3.2 光响应与气体交换参数测量
于采收期前30 d, 选择晴朗天气的9: 00—11: 00, 使用Li6400(LI-COR, USA)对最长枝条上顶端的成熟叶片进行光响应曲线参数测量, 每处理重复3盆。测量时, 叶室内相对湿度61.48%—65.04%, 叶室内温度为24.22—25.36 ℃, 光响应曲线的CO2浓度为380 μmol·mol–1, 光强分别设定为2000、1 800、1 500、1 200、1 000、800、600、400、200、150、100、70、50、20和0 μmol·m–2·s–1。根据直角双曲线修正模型, 计算光补偿点(LCP)、暗呼吸绝对值(Rd)、饱和光照强度(LSP)和最大净光合速率(Pmax)和表观量子效率(AQY)等光响应曲线的参数。在叶室内的CO2浓度和光合有效辐射分别为380 μmol·mol–1与1000 μmol·m–2·S–1时, 测定净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和气孔导度(Gs)[10]。
3.3 生长分析参数和叶绿素含量的测定
测量光合生理参数后, 调查测量4种光环境下延胡索的叶面积。地上部茎叶和地下部块茎(包括须根)带回实验室, 以烘干法80 ℃条件下测得干重。以上测定重复3次。同时, 采集4种不同光环境的延胡索鲜叶样品于实验室测定Chl a和Chl b, 用95%乙醇提取, 使用分光光度计分别在665 nm和649 nm下测定含量[11], 重复3次。
3.4 统计分析
采用Microsoft Excel 2007和SPSS 15软件对数据进行统计分析, 采用LSD法进行多重比较。
4 结果与分析
延胡索在大田高光照条件的光补偿点为44.91± 10.44 µmol·m–2·s–1(表1), 随着有效光辐射的减弱, 其光补偿点值呈下降趋势, 中低光照和弱光条件的光补偿点显著降低(<0.05); 弱光条件的暗呼吸绝对值最大, 与其他三个处理有显著差异(<0.05), 说明黑网的弱光环境已对其产生了胁迫作用; 饱和光照强度的值随着有效光辐射的减弱而减小, 且中低光照和弱光条件的值与高光照条件的值有显著差异(<0.05); 中高光照条件的平均最大净光合速率最高(10.95±2.76 µmol·m–2·s–1), 弱光条件下最小(4.29±0.85 µmol·m–2·s–1), 弱光条件的值与其他三个处理有显著差异(<0.05); 表观量子效率的变化趋势与光补偿点、饱和光照强度类似, 随着有效光辐射的减弱而降低, 且弱光条件的值与其他三个处理有显著差异。中高光照条件下, 延胡索的光响应曲线参数与高光照条件的差异并不显著(>0.05), 而黑网的弱光条件下, 各项光响应曲线参数明显下降, 表明黑网弱光对延胡索的光合作用有显著的限制。遮阴后, 延胡索的光补偿点、饱和光照强度和最大净光合速率均显著降低, 说明延胡索的光合机构对弱光环境有明显的响应, 以适应弱光环境; 在中低光照下, 延胡索的5个光响应特征参数均有向耐阴植物变化的趋势, 其通过降低暗呼吸速率和光补偿点等生理行为, 减少呼吸消耗、增强对低光的利用能力来维持碳收获, 从而保证自身的生长。

表1 延胡索不同光环境下的光响应曲线参数
注: 表格数据为平均值±标准差; 不同小写字母表示差异显著(<0.05)。下同。
延胡索在高光照条件下的净光合速率为8.47± 1.35 µmol·m–2·s–1(表2), 中高光照与中低光照条件下, 其均值有略微的下降, 但不显著(p<0.05), 弱光条件下则显著下降(<0.05, 4.96±1.15 µmol·m–2·s–1); 延胡索在不同光环境中的蒸腾速率、气孔导度的变化趋势与净光合速率的变化类似, 其均值都随着有效光辐射的减弱而下降, 中低光照、弱光条件的蒸腾速率与高光照有显著差异(<0.05), 但四种不同光环境下的气孔导度没有显著差异(>0.05); 延胡索胞间CO2浓度表现为则与上述三者相反。表明相对于高光照条件而言, 延胡索亦可适应中高光照的光环境, 而黑网的弱光环境显著降低了其净光合速率和蒸腾速率, 提升了胞间CO2浓度, 限制了其气体交换。
延胡索Chl a含量以中高光照条件最高(图1), 为2.27±0.34 mg·g–1, 高光照条件其次, 然后随着有效光辐射的降低, 其均值呈下降趋势, 高光照、中高光照和中低光照三者的Chl a含量差异不显著(>0.05), 而弱光条件Chl a与前三者均有显著差异(<0.05); 四种条件下的Chl b含量差异均不显著(<0.05), 但其均值以中高光照条件的最高, 中低光照条件次之; 随着有效辐射的下降, Chl a+Chl b含量先升后降, 中高光照条件的延胡索植株Chl a+Chl b含量最高, 弱光条件的最低, 弱光条件下的Chl a+Chl b含量与其他三种条件呈显著性差异(<0.05), 高光照、中高光照和中低光照之间差异性不显著(>0.05); 高光照条件的Chl a/Chl b值最高, 并随着有效光辐射的下降, Chl a/Chl b值亦下降, 四种条件Chl a/Chl b值互相之间均有显著性差异(<0.05)。
延胡索在中高光照条件下的叶面积均值最大(表3), 高光照条件次之, 弱光条件最小, 但高光照、中高光照和中低光照三种条件的叶面积差异不显著(>0.05), 弱光条件的叶面积与前三者有显著性差异(<0.05); 弱光条件下的叶生物量与前三种条件有显著相差异(<0.05), 且随着有效光辐射的减弱, 其均值呈下降趋势; 根生物量的表现与叶生物量趋同, 且中低光照条件的根生物量与高光照、中高光照两种条件均有显著性差异(<0.05); 四种条件的比叶面积均没有差异(>0.05), 但其均值表明, 中低光照条件下的光照较弱, 叶片较薄, 而有效光辐射最弱的弱光条件, 其比叶面积均值却与有效光辐射较强的中高光照条件相似, 应该与弱光抑制延胡索茎叶营养器官发育有关; 高光照、中高光照和中低光照三者的根冠比差异不显著(>0.05), 弱光条件的根冠比则与前三者有显著差异(<0.05), 四种条件的根冠比均值以中高光照的最高, 高光照次之, 表明中高光照处理可以一定程度地促进延胡索的根系生长, 提升根冠比。

表2 延胡索不同光环境下的气体交换参数
注: 表格数据为平均值±标准差; 不同小写字母表示差异显著(<0.05)。

图1 不同光环境中延胡索的叶绿素含量
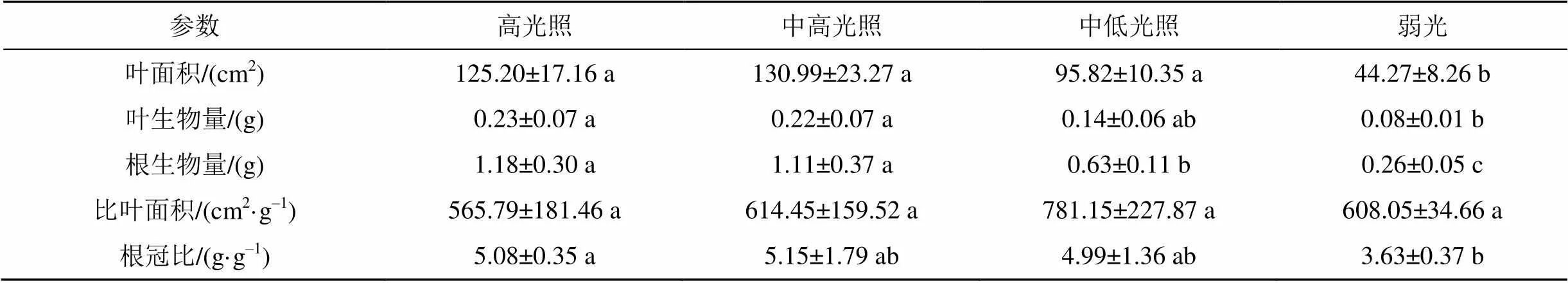
表3 延胡索不同光环境下的生长分析参数
注: 表格数据为平均值±标准差; 不同小写字母表示差异显著(<0.05)。
5 讨论
5.1 延胡索光合特征对光照的响应
在植物自然生境中, 光资源在时空上分布常表现出强烈的异质性[4], 超出植物适应范围的强光或弱光会抑制其正常生长发育[1, 3]。本试验表明, 就光响应参数而言, 在高光照和中高光照的条件下, 早春植物延胡索对两种光环境表现差异不显著, 在暗呼吸速率和最大净光合速率方面, 中高光照条件的均值甚至超过高光照条件, 说明高光照条件对于延胡索而言并不合适。弱光条件下的饱和光照强度、最大净光合速率和表观量子效率分别为高光照的62.67%、43.25%和58.42%, 其所有光响应参数都不同程度的下降或显著下降, 这与邹长明等[1]进行四种豆科作物的研究结果类似, 说明延胡索对弱光环境有明显地响应。同时, 亦表明延胡索对光辐射的适应范围较宽, 既能在高光照条件下不受光抑制, 也能适应弱光环境并维持碳水化合物的积累, 具备一定的耐阴能力。张云等[12]的研究表明, 植物通过降低光饱和点、光补偿点和暗呼吸速率, 提高表观量子效率等方式以适应弱光环境。但本试验在弱光条件下的表观量子效率却呈现显著下降, 这与延胡索在13.8%全光照的弱光环境中, 地上茎不能正常发育有关。
多数研究表明, 遮阴条件或林下植物的净光合速率、气孔导度和蒸腾速率总体均低于大田或林外高光照[4, 13]。本试验表明, 随着有效光辐射的减弱, 延胡索的净光合速率、蒸腾速率和气孔导度亦有不同程度的下降, 亦佐证了其光响应特征参数的变化趋势。同时, 延胡索在弱光环境中气孔导度较低, 叶片气孔开度下降, 气体交换能力较差, 单位时间内CO2分子进入量较少, 减少了水分子的反向逸出量, 胞间CO2浓度的升高也说明了这一现象, 极有可能是弱光条件抑制植物叶片的气孔的形成及生长, 叶片气孔数变少, 叶片气孔指数及气孔比率都有所降低造成的[6]。
植物可自身调节光合色素含量以适应不同的光环境[1, 13]。由于Chl b的功能主要是收集光能, 因此在弱光条件下, Chl b的含量显得尤为重要, Tanaka等[14]认为, 弱光条件下, 叶片的叶绿素酸脂a氧化酶的表达量和活性增加, 催化Ch1 a向Ch1 b转化, 这可能是弱光环境中叶片Ch1 b含量增加的主要原因。本试验中, 延胡索在遮阴后, 叶片中的Chl a、Chl b及其Chl a+Chl b皆随着有效光辐射的下降呈现了先升后降的趋势; 而Chl a/Chl b值是植物耐阴性能的重要指标之一[15], Chl a/Chl b值降低有利于充分利用弱光, 亦是耐阴植物对弱光环境的响应[12], 延胡索Chl a/Chl b值在遮阴(中高光照和中低光照)后显著降低, 表现出其耐阴的性能, 而弱光条件下Chl a/Chl b值显著下降的原因是遮阴弱光条件导致延胡索的光合产物供应不足, 妨碍了叶绿素的合成, 使其光合能力和生长发育严重受阻。这与弱光胁迫对七叶一枝花()光合特性有较显著的影响的研究结果类似[16], 表明延胡索具有一定的耐阴能力, 但并不适合长期的弱光条件。
张建新等[17]认为, 光饱和点<460 µmol·m–2·s–1且光补偿点<46 µmol·m–2·s–1的是耐阴植物, 否则即为喜光植物; 同时还认为, 当Chl a+Chl b>3.8 mg·g–1、Chl a/ Chl b<3.2时, 可以此为依据判断是否为耐阴植物。本试验延胡索的光饱和点远高于上述临界值, 而光补偿点却略小于临界值, Chl a+Chl b最大值为中高光照条件下的2.78±0.50 mg·g–1, Chl a/Chl b在中低光照和弱光条件下可以满足耐阴植物的临界值范畴, 造成这种情况的原因可能是: 一、取样和统计可能存在着一定误差; 二、以光饱和点和光补偿点等原本依赖于经验模型计算得到的文献资料数据来区分耐阴植物和喜光植物有一定限制性; 三、早春植物利用林下光辐射主要基于物种的时间生态位, 早春植物可能是一类较为特殊的植物类群, 延胡索既不是典型的阳性植物也不是典型的阴性植物, 其耐阴能力介于阳性植物和阴性植物之间, 弱光下的延胡索光合机构可能已向耐阴植物转变。
5.2 光照对延胡索生物量分配的影响
植物可通过改变表型可塑性的形式适应不同生存环境[18]。植物因光资源的异质性而发生根、茎及叶片形态结构、生物量分配格局的变化, 对于增强其生存能力和竞争能力极为重要[19]。研究发现, 有效光辐射过弱会影响七叶一枝花光合的各项参数, 40%或60%的弱光条件时达到最高, 超过80%后光合参数指标迅速下降[16]。在弱光环境下, 植物的根冠比通常会降低, 比叶面积上升[20]。本试验发现, 相对于高光照条件, 弱光条件下延胡索地上部和地下部的生物量呈现显著下降的变化, 主要表现在, 其叶生物量、根生物量分别仅为高光照条件下的34.78%和22.03%, 弱光条件下的根系几乎没有生长, 表明延胡索虽然具有一定的耐阴能力, 但弱光环境中对于非耐阴植物而言是一种光胁迫, 弱光条件下延胡索植株未形成花和果等器官, 这与观察到的林下郁闭度较高的野生延胡索种群开花较晚, 结实量较少相符; 而中高光照遮阴可以使根冠比略微上升, 表明适当遮阴对于延胡索根系生物量的积累有益; 而随着有效光辐射的下降, 比叶面积先升后降, 在13.8%高光照的弱光条件下, 其光合器官构型能力比高光照条件小, 但单位面积的干重优于中高光照和中低光照条件下的植株, 即弱光条件下的延胡索优先于叶片的形态构成, 而不再倾向于扩大叶面积捕获更多的光量子。
5.3 林下仿野生栽培延胡索的建议
林药复合生态系统可改善不同目标物种的群落结构, 增强种群抗逆性, 减少化肥、农药的施用量, 提高光能利用率、资源利用率和土地产出率[8, 16, 21]。延胡索为典型的早春植物, 株型不高, 生长期短, 其生长发育期可分为萌芽发根、地下茎生长、出苗及地上部分生长和块茎生长4个阶段。延胡索出苗后, 利用早春时间(2—4月)疏林下空隙光斑进行光合作用, 叶生长和根系生长的适宜平均温度为10 ℃—18 ℃, 待乔灌层树种抽枝散叶时便开花结实, 当均温升至22 ℃时, 叶片开始出现枯焦, 25 ℃时则茎叶出现枯萎直到死亡, 结束其整个生活史。根据延胡索在不同光环境中的光合与生长的表现: 适当遮阴(中高光照)时, 其光响应特征参数并没有显著下降, 表明延胡索不同程度地向耐阴植物变化, 仍能维持较高的干物质积累效率, 完成生活史, 且有益于根系生物量的积累, 适宜在疏林间作套种; 试验中发现, 中低光照遮阴对延胡索已经出现一定的抑制作用, 使其开花数量减少, 种子产量降低; 而弱光条件对延胡索是一种胁迫, 遮阴后其生长发育严重受阻, 干物质积累效率显著降低且不能开花结籽。因此, 我们建议: ⑴高光照条件下, 3-5月延胡索地上部分生长发育时, 可以进行80%全光照的遮阴处理; ⑵野外疏林下进行仿野生延胡索栽培时, 当乔冠层的遮阴度(郁闭度)过高时, 可适当采取间伐修枝措施或者开辟林窗改善林下光环境, 提升林下延胡索的光合作用能力和光斑利用效率; ⑶4月进行遮阴处理(80%全光照)可以延长林下延胡索地上部分生存的时间, 合理配置其光生态位和时间生态位以达到预期的经济效益。
[1] 邹长明, 王允青, 刘英, 等. 四种豆科作物的光合生理和生长发育对弱光的响应[J]. 植物生态学报, 2015, 39(9): 909–916.
[2] KNOX J, MORRIS J, HESS T. Identifying future risks to UK agricultural crop production: Putting climate change in context[J]. Outlook on Agriculture, 2010, 39(39): 249–256.
[3] 欧建德, 吴志庄. 林下套种及坡位和弱光环境对南方红豆杉人工林早期生长及林分分化的影响[J]. 东北林业大学学报, 2016, 44(10): 12–16.
[4] BLOOR J M G, GRUBB P J. Growth and mortality in high and low light: Trends among 15 shade-tolerant tropical rain forest tree species[J]. Journal of Ecology, 2003, 91(1): 77–85.
[5] 江泽慧, 吴泽民, 何云核, 等. 绞股蓝的林下栽培[J]. 植物资源与环境学报, 1995, 4(1): 13–16.
[6] 吴德成, 牟兆军, 柏松林, 等. 林下与效应带种植人参环境因子的动态变化[J]. 植物研究, 1995, 15(1): 118–128.
[7] GAUDIO N, BALANDIER P, DUMAS Y, et al. Growth and morphology of three forest understory species (,and) according to light availability[J]. Forest Ecology and Management, 2011, 261: 489–498.
[8] 毛世忠, 赵博, 蒋小华, 等. 林下不同光照条件对走马胎苗木生长及光合特性的影响[J]. 西北林学院学报, 2016, 31(1): 21–24.
[9] LI Weicheng, SHENG Haiyan. Effects of bamboo charcoal-based biofertilizer on wine bamboo sap yield and nutrient composition[J]. Journal of Forest Research. DOI 10.1007/s11676–017–0516–2.
[10] 李伟成, 盛海燕, 高贵宾, 等. 高温干旱对髯毛箬竹的叶和细根的生理生态影响[J]. 环境科学研究, 2017, 30(12): 1908–1918.
[11] 王学奎. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社, 2006: 134–139.
[12] 张云, 夏国华, 马凯, 等. 遮阴对堇叶紫金牛光合特性和叶绿素荧光参数的影响[J]. 应用生态学报, 2014, 25(7): 1940–1948.
[13] 刘延惠, 姜霞, 丁访军, 等. 头花蓼光合特性及叶绿素荧光对林下光照生境变化的响应[J]. 水土保持学报, 2017, 31(6): 345–352.
[14] TANAKA R, TANAKA A. Effects of chlorophyllide a, oxygenase overexpression on light acclimation in[J]. Photosynthesis Research, 2005, 85(3): 327–340.
[15] URBAS P, ZOBEL K. Adaptive and inevitable morphological plasticity of three herbaceous species in a multi-species community: field experiment with manipulated nutrients and light[J]. Acta Oecologic, 2000, 21(2): 139–147.
[16] 龚范武, 龚舟, 彭静, 等. 不同林下七叶一枝花栽培技术研究[J]. 中国农学通报, 2015, 31(13): 155–158.
[17] 张建新, 颜赞, 方炎明. 遮光对臭牡丹生长和光合特性的影响[J]. 植物资源与环境学报, 2013, 22(1): 88–93.
[18] 王浩, 李德志, 程立丽, 等. 光资源强度变化对结缕草植株克隆生长特征的影响[J]. 生态科学, 2014, 33(4): 664–671.
[19] 李良, 夏富才, 孙越, 等. 阔叶红松林下早春植物生物量分配[J]. 北京林业大学学报, 2017, 34(1): 34–42.
[20] OSADA N, TAKEDA H, KITAJIMA K, et a1. Functional correlates of leaf demographic response to gap release in saplings of a shade-tolerant tree,[J]. Oecologia, 2003, 137(2): 181–187.
[21] 卢玉鹏, 许纪元, 张晓曦, 等. 林下药用植物淋出物对红桦和杜仲枯落物分解及土壤酶活性的影响[J]. 植物生态学报, 2017, 41(6): 639–649.
Responses of photosynthesis and growth plasticity ofto light
SHENG Haiyan1, LI Weicheng2, 3, *
1. Hangzhou Environmental Protection Science Institute, Hangzhou 310005, China 2. China National Bamboo Research Center/Key Laboratory of High Efficient Processing of Bamboo of Zhejiang Province, Hangzhou 310012, China 3. College of Life Sciences, Southwest Forestry University, Kunming 650224, China
Medicinal plant-based agroforestry systems have considerable economic benefits. In the present study, the changes in the photosynthesis and growth of the early spring plant,, under different light conditions were analyzed. This was done to provide a scientific basis for understory tending, canopy thinning, and development of medicinal plant-based agroforestry systems using this species. It was found thatadapted to shaded and low-light environments by decreasing its light saturation point, light compensation point, and dark respiration rate, and increasing its apparent quantum yield. Differences in the performance ofunder full- and medium-light conditions were not significant. Particularly, the mean values for dark respiration rate and maximum net photosynthetic rate under medium-light conditions were higher than those under full-light conditions. The saturation light intensity, maximum net photosynthetic rate, and apparent quantum yield under low-light conditions were 62.67%, 43.25%, and 58.42% of the values under full-light conditions, respectively. The net photosynthetic rate, stomatal conductance, and transpiration rate under shaded conditions were generally lower than the corresponding values under full-light conditions. After shading, Chl a, Chl b, and Chl a + Chl b levels in the leaves ofinitially increased and subsequently decreased as photosynthetically active radiation decreased. Under medium-high- and medium-low-light conditions, the Chl a: Chl b ratio significantly decreased. This demonstrates the shade tolerance of this species, whereas the significant decrease in the Chl a: Chl b ratio under low-light conditions is attributed to the insufficient supply of photosynthates and reduced chlorophyll synthesis caused by low light. The leaf and root biomasses ofunder low-light conditions were only 34.78% and 22.03% of the values obtained under full-light conditions, respectively. With a decrease in photosynthetically active radiation, the specific leaf area initially increased and subsequently decreased, indicating thatprioritizes the morphological construction of leaves over the expansion of leaf area to capture more photons under low-light conditions. The shade tolerance ofis intermediate between that of light-demanding plants and shade plants, andthe photosynthetic apparatus ofmay become similar to that of shade-tolerant plants under low-light conditions. Therefore,has a certain level of shade tolerance, but it is not well-suited for long-term exposure to low-light conditions. During the cultivation ofin wild woodlands with very high canopy closure, measures such as appropriate thinning and pruning, or creation of forest gaps, can improve understory light environment and enhance photosynthetic capacity and sunfleck utilization of understoryplants. These measures can then allow for reasonable allocation of light and temporal niches for the plants to achieve the expected economic benefits.
; early-spring plant; weak light; tree-herb plantations; ecophysiology
10.14108/j.cnki.1008-8873.2018.06.022
Q145
A
1008-8873(2018)06-168-07
2018-04-07;
2018-07-16
浙江省科技计划项目(2016F50008); 浙江省省院合作项目(2015SY06)
盛海燕(1978—), 女, 浙江临安人, 高级工程师, 主要从事生态学研究, E-mail: laohaishy@163.com
李伟成, 男, 博士, 副研究员, 主要从事生态学研究, E-mail: lee_caf@163.com
盛海燕, 李伟成. 延胡索光合与生长可塑性对光照的响应[J]. 生态科学, 2018, 37(6): 168-174.
SHENG Haiyan, LI Weichen. Responses of photosynthesis and growth plasticity ofto light[J]. Ecological Science, 2018, 37(6): 168-174.
