氯化钙处理对菠萝黑腐病的防控效果及机制分析
2019-01-09谷会贾志伟侯晓婉张鲁斌
谷会 贾志伟 侯晓婉 张鲁斌

摘 要 为了降低采后菠萝果实由黑腐病引起的腐烂损失,本文研究了氯化钙处理对黑腐病的防控效果及相关机制。结果表明,1%氯化钙处理对菠萝黑腐病有最好的抑制作用和防治效果。1%氯化钙处理对病菌菌丝生长影响不大,但可明显抑制病原菌的产孢;氯化钙处理明显降低发病果实中多聚半乳糖醛酸甲酯酶(PMG)、纤维素酶(CX)和β-葡萄糖苷酶(CB)的活性,但对多聚半乳糖醛酸酶(PG)活性变化没有明显影响;氯化钙处理明显提高贮藏中后期菠萝果实中过氧化氢(H2O2)的含量,明显提高抗病相关酶苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)、几丁质酶(CHI)和β-1,3-葡聚糖酶(GLU)的活性。说明氯化钙处理对黑腐病有较好的防控效果,一方面是由于其降低了病原菌的致病性,另一方面是诱导菠萝果实提高了抗病性。因此,适宜浓度的氯化钙处理为菠萝采后黑腐病的绿色防控提供了一条新的途径。
关键词 菠萝;黑腐病;氯化钙;防控效果
中图分类号 S436 文献标识码 A
Effects and Mechanism of Calcium Chloride Treatment on Reducing Pineapple Black Rot
GU Hui, JIA Zhiwei, HOU Xiaowan, ZHANG Lubin*
South Subtropical Crops Research Institute, Chinese Academy of Tropical Agricultural Sciences / Hainan Key Laboratory for Postharvest Physiology and Technology of Tropical Horticultural Products, Zhanjiang, Guangdong 524091, China
Abstract In order to reduce the loss of pineapple fruit caused by black rot after harvest, the effects of calcium chloride treatment on black rot and related mechanism were studied in this paper. The results showed that 1% calcium chloride treatment had the best inhibitory and control effect. Calcium chloride treatment (1%) had little effect on the growth of mycelial, but could significantly inhibit the sporulation of pathogens. Calcium chloride treatment significantly reduced the enzymatic activities of PMG, CX and CB in the treated fruits, but had no significant effect on the changes of PG activity. Calcium chloride treatment significantly increased the content of H2O2 during the middle and late storage, and significantly increased the activities of resistance-related enzymes such as PAL, PPO, CHI and GLU. These results showed that calcium chloride treatment displayed effects on inhibiting black rot probably due to it reduced the pathogenicity of pathogens and induced disease resistance of treated pineapple fruits. Therefore, the appropriate concentration of calcium chloride treatment would provide a new way for the green control of pineapple black rot after harvest.
Keywords pineapple; black rot; calcium chloride; control effect
DOI 10.3969/j.issn.1000-2561.2019.12.023
菠蘿黑腐病是一种真菌性病害,病原菌是奇异根串珠霉(Thielaviopsis paradoxa),黑腐病是采后菠萝果实主要的侵染性病害,在国内外菠萝种植地区均有发生,国内有的产区发病率高达40%,严重影响了菠萝的市场供应及加工生产[1]。前人研究表明[2],病原真菌通过向果实内分泌细胞壁降解酶、毒素、病原物激素来降解寄主的细胞壁和角质层,从而使病原真菌的侵入与扩展,而细胞壁降解酶是主要的致病因子。同样,细胞壁降解酶在菠萝黑腐病菌的致病过程中也起着很重要的作用,主要包括多聚半乳糖醛酸酶(PG)、多聚半乳糖醛酸甲酯酶(PMG)、果胶甲酯酶(PME)、纤维素酶(CX)和β-葡萄糖苷酶(CB)等。PG、PMG和PME在侵染前期降解细胞壁中果胶质使果实软化,CX和CB在侵染中后期降解细胞壁中的纤维素使果实细胞壁结构破坏,利于黑腐病菌的侵入与扩展。
化学杀菌剂是防治菠萝采后黑腐病有效措施,目前在生产上常用于防治菠萝果实黑腐病的杀菌剂有多菌灵、甲基托布津和苯菌灵等[3]。Wije singhe等[4]研究发现,用3 g/kg的苯菌灵浸果3 min、或用0.2 g/L的抑霉唑浸果1 min,对菠萝黑腐病防效明显,但长期使用化学杀菌剂一方面可导致病原菌产生抗药性,另一方面也存在食品安全隐患。Wilson等[5]用54 ℃热水处理菠萝3 min后,菠萝在10 ℃贮藏21 d后没有发现黑腐病,但由于热水处理程序繁琐,容易引起交叉感染,在生产上难以推广。也有研究者用拮抗木霉(Trichode rma asperellum)[6]来防治菠萝黑腐病,但由于拮抗菌对营养和环境条件的特殊要求以及在菠萝果实上难以定植的问题,拮抗菌在生产上大面积应用还有较大困难。
能诱导采后果蔬产生抗病性的因素包括生物激发子和非生物激发子,钙离子作为一种非生物激发子,对采后果实也有一定的诱导抗病效果,比较常见的保鲜剂是氯化钙。氯化钙处理成本低,使用安全方便,通过植物体内的生理生化反应来发挥作用,不会对环境造成污染,适合绿色农业的发展要求和标准[7]。關于钙处理在采后果蔬上的抗病性研究已有很多报道,Conway最早在1983年就研究发现,采后钙处理能提高苹果果实的诱导抗病性,钙真空渗透处理对苹果灰霉病有较好的防治效果[8]。之后分别在桃[9]、梨[10]、芒果[11]以及苹果[12-13]上均有报道。还有研究表明,钙处理对苹果炭疽菌的生长有直接的抑制作用[14-15]。但氯化钙处理是否可以提高采后菠萝果实的抗病性,是否对菠萝黑腐病菌生长有抑制作用还未见报道。所以本研究以此为切入点,研究氯化钙处理对菠萝黑腐病的防治效果,并测定氯化钙对黑腐病菌的抑制作用以及对其致病性的影响,同时还研究氯化钙处理对菠萝果实抗病性的影响,以期为菠萝黑腐病的绿色防控提供理论依据和应用参考。
1 材料与方法
1.1 材料
1.1.1 供试菠萝品种 菠萝品种为‘巴厘菠萝(Ananas comosus ‘Comtede Paris),采摘于广东省湛江市徐闻县菠萝实验基地,采摘时挑选果眼平整、果皮全绿的七成熟果实,立即运回南亚热带作物研究所实验室备用。
1.1.2 化学试剂 无水氯化钙(分析纯),天津市福晨化学试剂厂;蜗牛酶(分析纯),Sigma公司;昆布多糖(分析纯),Sigma公司;几丁质(分析纯),上海伊卡生物技术有限公司;N-乙酰氨基葡萄糖,Sigma公司;多聚半乳糖醛酸(分析纯),Biosharp公司;果胶质(分析纯),Biosharp公司;羧甲基纤维素钠(分析纯),天津市福晨化学试剂厂;D-半乳糖醛酸(分析纯),Biosharp公司;3,5-二硝基水杨酸(化学纯),国药集团化学试剂有限公司。
1.1.3 供试菌株 病原菌株为菠萝黑腐病菌(Thielaviopsis paradoxa),为南亚热带作物研究所采后病理实验室分离保存。
1.1.4 仪器与设备 UV-2700紫外可见分光光度计,日本岛津公司;Envolution RC型超速离心机,美国Thermo Fisher公司;MIR-553型低温恒温培养箱,日本Sanyo公司。
1.2 方法
1.2.1 氯化钙处理对菠萝黑腐病病斑直径抑制率的测定 氯化钙是一种无毒的绿色保鲜剂,对采后苹果、梨等果蔬有较好的防腐保鲜作用,本实验以氯化钙为保鲜防腐剂,开展菠萝黑腐病防治技术研究。首先将氯化钙配制成质量浓度分别为0.5%、1%、2%的水溶液,将菠萝果实分别在不同浓度的氯化钙保鲜液中浸泡2 min,取出自然晾干,实验以清水浸泡为对照,用PE袋敞口包装,置于常温25 ℃下贮藏12 h后,在菠萝果面中间接种浓度为1×106个/mL的黑腐病菌孢子悬浮液15 μL,置于低温15 ℃下贮藏。每个处理10个果实,每隔3 d调查一次病斑直径大小,并计算抑制率,重复3次。
1.2.2 氯化钙处理对菠萝黑腐病的防治效果 将菠萝果实在1%的氯化钙水溶液中浸泡5 min,以清水浸泡为对照,取出后自然晾干,用PE袋敞口包装,置于低温15 ℃下贮藏。每处理60个果实,每隔3 d取15个果实调查病情指数,并计算防治效果,重复3次。黑腐病的发病情况按果实纵剖面上病害面积大小分为0~5级,0级:无病害;1级:病害面积占剖面面积的10%以下;2级:病害面积占比11%~25%;3级:病害面积占比26%~50%;4级:病害面积占比51%~75%;5级:病害面积占比超过75%。
病情指数=Σ[(病害果数×病害级值)/总果数×最高病害级值]×100
防治效果=(对照病情指数–处理病情指数) /对照病情指数×100%
1.2.3 氯化钙对菠萝黑腐病菌菌丝生长和产孢影响 将氯化钙加入到定量灭菌的马铃薯葡萄糖琼脂培养基(PDA)中制备成0.5%、1%、2%的氯化钙含药PDA平板,从培养48 h的菌落边缘打取直径为5 mm的菌苔,分别接种到上述含药PDA平板中央,以不含药PDA平板作为对照,置于25 ℃下恒温培养,培养72 h后测量菌落的直径,并用血球计数板测定每个培养皿的产孢个数,3次重复。
1.2.4 菠萝果实中细胞壁降解酶活性的测定 挑选无病虫害、无机械损伤、大小基本一致的菠萝果实,在1%的氯化钙保鲜液中浸泡5 min,取出自然晾干,清水浸泡作为对照,在25 ℃下贮藏12 h后,针刺接种菠萝黑腐病菌,15 ℃下贮藏,每处理25个果,重复3次,分别在第0、3、6、9、12 d取5个果实进行取样。在果实病健交界处切取0.5 g果皮组织,用0.05 mol/L pH 8.0的Tris-HCl缓冲液(内含1%聚乙烯吡咯烷酮、10 mmol/L EDTA,1 mol/L NaCl)进行提取,在4 ℃、10000 r/min下离心25 min,取上清液作为待测酶液。菠萝果实内细胞壁降解酶PG、PMG、CX、CB酶活性的测定均采用3,5-二硝基水杨酸(DNS)比色法[16],其中PG和PMG酶活性测定用D-半乳糖醛酸制作标准曲线,以每小时催化底物释放出1 mg D-半乳糖醛酸的量为1个酶活单位U,酶活性以U/g 计算;CX和CB酶活性的测定用葡萄糖制作标准曲线,以每小时催化底物释放出1 mg葡萄糖的量为1个酶活单位U,酶活性以U/g计算,所有酶活性测定均3次重复。
1.2.5 菠萝果实过氧化氢含量的测定 菠萝果实的前处理和取样方法同1.2.4。取1 g菠萝果皮,用0.05 mol/L pH 7.0的磷酸缓冲液提取,粗酶液取样量为0.5 mL,采用南京建成生物工程研究所生产的过氧化氢试剂盒测定,过氧化氢含量以μmol/g计算,3次重复。
1.2.6 菠萝果实抗病相关酶活性的测定 菠萝果实的前处理和取样方法同1.2.4。苯丙氨酸解氨酶(PAL)活性测定参考吴芳芳等[17]的方法,取病健交界处的菠萝组织1 g,提取介质为0.05 mol/L pH 8.8硼酸缓冲液(内含1%聚乙烯吡咯烷酮、5 mmol/L巯基乙醇、1 mmol/L EDTA),于290 nm波长下测OD值,每分钟改变0.01为一个酶活单位U,以U/g计算。多酚氧化酶(PPO)活性测定参考Zhang等[18]的方法,在病健交界处取1 g菠萝果皮,提取介质为0.05 mol/L pH 6.8 磷酸缓冲液,于398 nm波长下测OD值,每分钟改变0.01为一个酶活单位U,以U/g计算。几丁质酶(CHI)和β-1,3-葡聚糖酶(GLU)粗酶液的制备参考杨冬平等[19]的方法,在病健交界处取0.5 g菠萝果皮,用0.1 mol/L pH 5.0的乙酸钠缓冲液(内含5 mmol/L巯基乙醇、1 mmol/L EDTA)提取,用提取液在4 ℃下透析过夜,离心取上清液即为粗酶液。CHI酶活性的测定参照马桂珍等[20]的方法,用N-乙酰氨基葡萄糖制作标准曲线,以每分钟产生1 μg N-乙酰氨基葡萄糖的量为1个酶活单位U,以U/g计算,重复3次。GLU酶活性的测定参照李世贵等[21]的方法,用葡萄糖制作标准曲线,1个酶活单位U定义为每分钟从昆布多糖中释放1 μg葡萄糖所需的酶量,以U/g计算,重复3次。
1.3 数据处理
采用Excel 2010软件统计所有数据,计算均值和标准误并制图;数据的方差分析(ANOVA)采用SPSS 22.0软件,利用Duncans多重比较对数据的显著性差异进行分析,P<0.05表示差异显著。
2 结果与分析
2.1 氯化钙处理对菠萝黑腐病病斑直径的抑制率
研究结果表明,菠萝果实在15 ℃下贮藏3 d后,氯化钙处理和对照的病斑直径无显著性差异,但贮藏6、9、12 d后,0.5%、1%、2%氯化钙处理的病斑直径明显小于对照,表现出较好的抑制作用(图1)。其中1%的氯化钙表现出最好的抑制作用,贮藏6、9、12 d后的病斑直径分别为39.9、69.4、95.0 mm,抑制率分别为17.9%、14.8%、12.3%。
2.2 氯化钙处理对菠萝黑腐病的防治效果
图2结果表明,菠萝果实在15 ℃下贮藏3 d后开始发病,1%氯化钙处理和对照的病情指数无显著性差异,但贮藏6、9、12 d后,对照的病情指数分别为29.3、53.3、86.7,而氯化钙处理的病情指数为24.0、44.0、72.0,明显小于对照。贮藏6、9、12 d后,1%氯化钙处理的防治效果分别为18.1%、17.4%、16.9%。
2.3 氯化钙对菠萝黑腐病菌菌丝生长和产孢影响
从表1可以看出,病菌在含不同浓度氯化钙的培养基上生长72 h后的菌落直径和对照无显著性差异,表明氯化钙对离体菌丝生长没有明显的抑制作用;而在含有1%、2%氯化钙的培养基上生长72 h后的产孢量分别为0.8×106个/皿、0.7×106个/皿,明显小于对照的产孢量,而且浓度越大产孢越少,说明氯化钙对黑腐病菌的产孢有一定的抑制作用。
2.4 氯化钙处理对菠萝果实中细胞壁降解酶活性的影响
从图3A可以看出,氯化钙处理对菠萝果实PG酶活性的影响没有明显的规律,对照菠萝果实的PG酶活性在2.0 U/g左右呈上下波动趋势,在贮藏9 d后达到最高2.4 U/g,而氯化钙处理后的PG酶活性也是在2.0 U/g左右呈上下波动趋势,在贮藏6 d后反而会提高PG酶的活性,12 d后和对照没有显著性差异。
从图3B、图3C和圖3D可以看出,对照菠萝果实中PMG、CX和CB酶活性在贮藏9 d前呈上升趋势,在贮藏9 d后达到最大值2.1、1.5、1.5 U/g,之后缓慢下降,贮藏12 d后的酶活性分别为2.0、1.6、1.3 U/g,明显高于氯化钙处理。而氯化钙处理的菠萝果实中PMG、CX和CB酶活性在贮藏6 d前上升不明显,贮藏6 d后呈快速上升趋势,12 d后分别达到最高1.9、1.4、1.2 U/g。氯化钙处理明显降低了发病菠萝果实中PMG、CX和CB的酶活性,其中PMG和CX酶活性的降低程度在贮藏6 d时达最大,分别为0.38、0.35 U/g,CB酶活性的降低程度在贮藏9 d时达最大0.38 U/g。
2.5 氯化钙处理对菠萝果实过氧化氢含量的影响
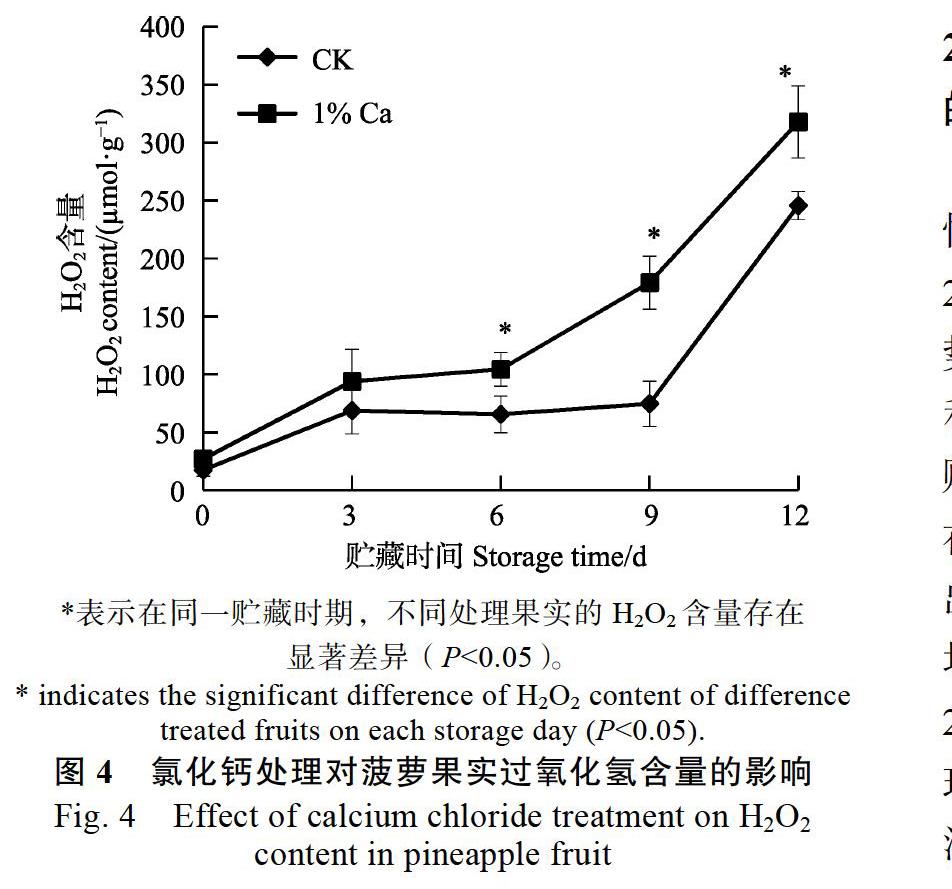
从图4可以看出,氯化钙处理和对照的菠萝果实中过氧化氢含量均呈上升趋势,氯化钙处理的菠萝在贮藏6 d后过氧化氢含量迅速上升,12 d
时达到最高值317.9 μmol/g,对照菠萝在贮藏9 d后过氧化氢含量迅速上升,12 d时达到最高值245.8 μmol/g。在贮藏3 d前氯化钙处理和对照菠萝果实中过氧化氢含量基本无显著性差异,在贮藏6、9、12 d后氯化钙处理明显高于对照。
2.6 氯化钙处理对菠萝果实抗病相关酶活性的影响
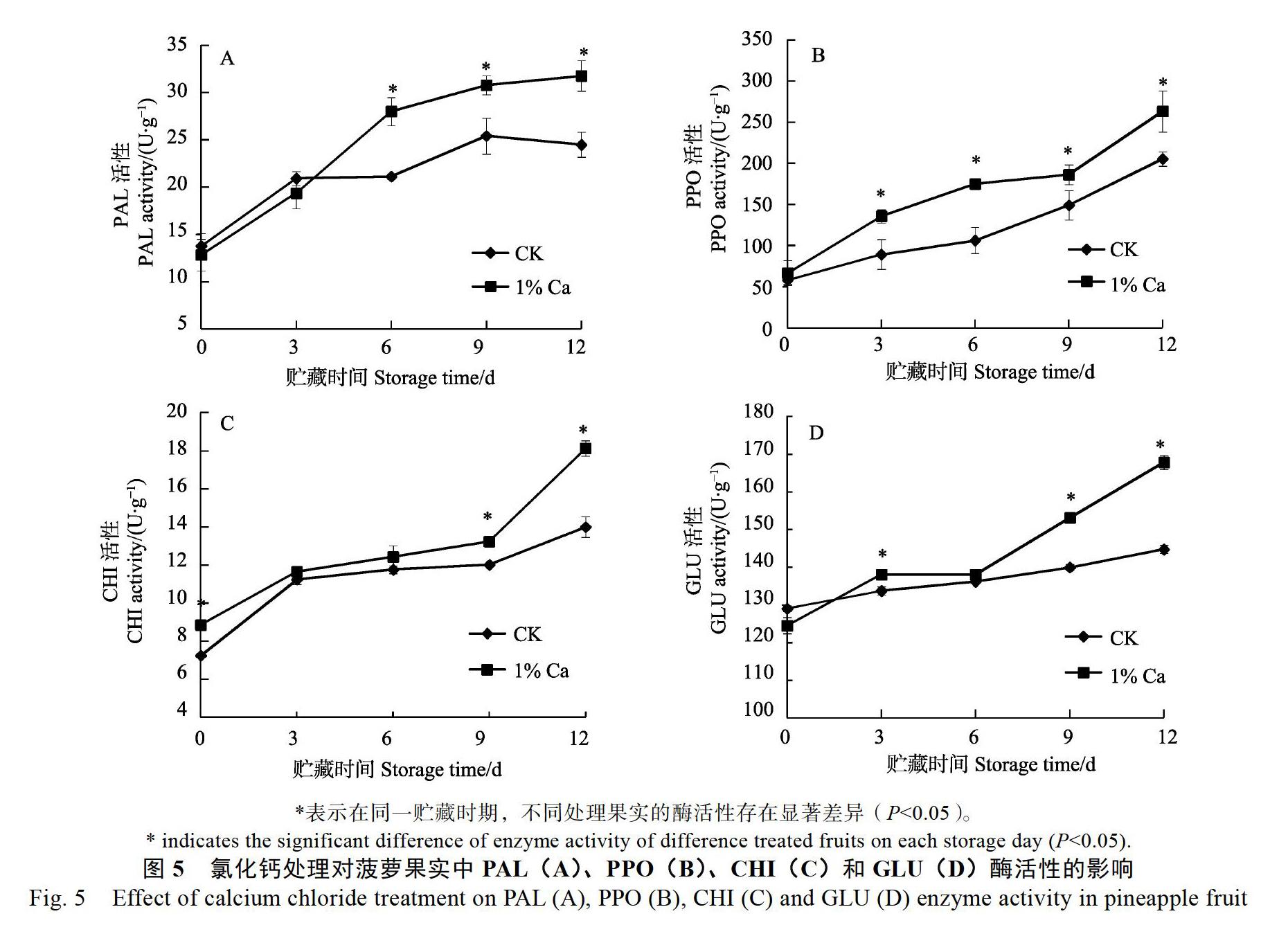
从图5A可知,对照菠萝果实中的PAL酶活性呈先上升后下降趋势,贮藏9 d时达到最大值25.4 U/g,而氯化钙处理在贮藏中一直呈上升趋势,到21 d时达到最大值31.8 U/g。氯化钙处理和对照的PAL酶活性在贮藏3 d前无显著性差异,贮藏6 d后到12 d,PAL酶活性明显高于对照,在贮藏12 d时比对照高7.3 U/g。从图5B可以看出,氯化钙处理和对照的菠萝果实中PPO酶活性均呈上升趋势,在贮藏12 d后分别达到最高值263.3、205.5 U/g。在整个贮藏过程中,氯化钙处理的菠萝果实中PPO酶活性均明显高于对照的酶活性。从图5C可知,氯化钙处理和对照的
菠萝果实中CHI酶活性在整个贮藏期间均呈上升趋势,贮藏9 d后上升较快,到12 d时达到最大,分别为18.1、14.0 U/g,贮藏9 d后氯化钙处理的CHI酶活明显高于对照。从图5D可以看出,氯化钙处理和对照的菠萝果实中GLU酶活性在整个贮藏期间均呈上升趋势,贮藏6 d时氯化钙处理和对照无显著性差异,贮藏6 d以后,氯化钙处理的GLU酶活迅速上升,贮藏9、12 d时的酶活性分别为153.2、167.8 U/g,而对照分别为140.0、144.7 U/g,氯化钙处理明显高于对照。
3 讨论
不同浓度的氯化钙处理对菠萝黑腐病均有一定的抑制作用,其中1%氯化钙表现出较好的抑制作用和防治效果,贮藏6、9、12 d后的抑制率分别为17.9%、14.8%、12.3%,防治效果分别为18.1%、17.4%、16.9%。其中贮藏6 d时的抑制作用和防治效果最好,之后呈下降趋势,这与病原菌的致病性以及菠萝果实的抗病性都有关系。贮藏6 d时防治效果最好,主要是由于氯化钙处理明显降低了菠萝果实中细胞壁降解酶的活性,而此阶段菠萝果实的诱导抗病效果不明显。而贮藏6 d以后防治效果呈下降趋势,主要是由于氯化钙对黑腐病菌的抑菌效果减弱,细胞壁降解酶的含量迅速提高,此阶段的防治效果主要来自氯化钙处理对菠萝果实的诱导抗病性。
氯化钙处理降低菠萝果实中细胞壁降解酶的活性,主要有两方面原因:一是由于氯化钙对黑腐病菌生长产孢的直接抑制作用,从而降低了病原菌分泌的细胞壁降解酶。关于这方面的研究已有相关报道,Chardonnet等[22]研究表明,钙处理可以降低真菌病害的侵染率是由于其对病原菌的直接抑制作用,包括抑制真菌的菌丝生长,孢子萌发和芽管伸长等。吴芳芳等[14]研究认为,钙盐处理可以明显降低苹果胶孢炭疽菌菌丝体干重和孢子萌发率。而本研究结果发现,氯化钙对黑腐病菌的菌丝生长快慢虽没有明显的抑制作用,但对黑腐病菌的产孢有较强的抑制作用,由于病原菌的生长一部分是来源于菌丝体营养生长,另一部分是来源于生殖生长,说明氯化钙可以降低病原菌的生长总量,从而减弱了黑腐病菌的致病性。另一方面,钙离子不但具有维持果实细胞壁和细胞膜结构和功能的作用,从而增强细胞壁对细胞壁降解酶的抵抗性;而且,氯化钙处理还可以降低果实自身产生的细胞壁降解酶活性。关于这方面的报道也有很多,Ortiz等[23]研究表明,钙处理可以减少苹果采后硬度的下降与果实中PG、PME 和果胶酸裂解酶(PL)活性的下降密切相关。张强等[24]研究发现氯化钙处理可以明显降低伽师瓜果胶酶PG、PME和PL的活性及其基因表达,延缓果实的软化。Ranjbar等[25]研究表明纳米钙处理可以明显降低苹果PG和PME酶的活性。另外,在柑橘[26]、番茄[27]、草莓[28]等其他水果上也有报道。而本研究结果发现,氯化钙处理对于菠萝PG酶的活性没有明显的抑制作用,但可以明显降低贮藏中后期菠萝果实中的PMG、CX和CB酶活性,这是由于氯化钙降低了病原菌分泌的PMG、CX和CB酶活性,也可能是降低了果实自身产生的细胞壁降解酶PMG、CX和CB的活性。
可以提高果实抗病性相关的酶主要有PAL、PPO、POD、病程相关蛋白等[29],PAL是果实抗病物质生成途径苯丙烷类代谢途径的关键酶和限速酶,有助于酚类物质的聚集,木质素的积累,植保素的合成等,PPO可氧化酚类化合物形成对病原菌更强毒性的醌类衍生物,POD参与抗病相关物质木质素与酚类的合成,病程相关蛋白(PR蛋白)的积累是果实获得系统抗性的重要原因,其中CHI和GLU是两种重要的PR蛋白,由于这两种酶可以降解病原菌细胞壁,所以其酶活水平可以直接证明果实抗病能力。关于钙处理可提高采后果蔬抗病相关酶的报道已有很多,Zhao等[13]研究发现,5%的氯化钙处理可提高苹果对炭疽病的抗性,与提高抗病相关酶PAL和POD的活性有关。Yu等[10]研究发现,氯化钙处理能够诱导梨果实抗病相关酶POD活性上升,从而提高梨对青霉病和灰霉病的抗病性。陈敏等[11]研究发现,40 g/L氯化钙与病原菌孢子处理间隔24 h,可显著提高其对芒果炭疽病的抑制效果,说明氯化钙能够诱导芒果果实抗病相关酶PAL、POD和PPO活性的上升,从而使其产生抗病性。同样,本实验研究结果也发现,氯化钙处理可以提高菠萝果实贮藏中后期的抗病相关酶PAL、PPO、CHI、GLU的活性,此阶段氯化钙对黑腐病的防治效果主要来自其对菠萝果实的诱导抗病性。
H2O2是典型的活性氧自由基,病原菌侵染和激发子均可诱导植物体内H2O2的产生,其直接作用就是抑制病原菌孢子萌发甚至杀死病原物,活性氧爆发是植物做出的快速防御反应之一[30];另一方面,H2O2还是植物防御反应中一个重要的信号物质,可作为植物的第二信使,在信号转导中调控下游信号,进而激活和调控植物体内各种胁迫相关基因,并将信号最终放大为蛋白的翻译表达,在植物体中即为产生各种抗病相关物质[31]。前人研究表明,不同的非生物激发子比如β-氨基丁酸[32-33],苯并噻二唑[34]均可诱导果实内H2O2大量生成,进而调控病程相关蛋白CHI和GLU的翻譯表达,提高果实的抗病性。本研究结果发现,钙离子作为一种非生物激发子,也可诱导菠萝果实内H2O2大量生成,尤其在贮藏后期产生活性氧爆发后,氯化钙处理的H2O2含量明显高于仅有损伤接种处理的对照,而在此阶段,氯化钙处理的菠萝果实CHI和GLU酶活性也明显高于对照。
总之,氯化钙处理对菠萝采后黑腐病有较好的防治效果,其中1%氯化钙处理的防治效果最好,最高为18.1%。氯化钙处理对黑腐病的防治机制主要有两方面:其一是氯化钙对黑腐病菌的产孢有直接的抑制作用,进而降低了菠萝果实中细胞壁降解酶的活性,延缓了果实的软化腐烂;其二是氯化钙处理对采后菠萝果实有一定的诱导抗病效果,主要表现为可以提高果实抗病性相关酶PAL、PPO、CHI、GLU的活性,可以提高贮藏后期果实中H2O2的含量,而H2O2含量的提高可以调控病程相关蛋白CHI和GLU的翻译表达,进而提高果实的抗病性。而信号物质H2O2如何调控菠萝果实内各种抗病基因的表达还有待进一步研究。
参考文献
谷 会, 朱世江, 詹儒林, 等. 菠萝黑腐病菌(Thielavi opsis paradoxa)生物学特性及其对杀菌剂敏感性测定[J]. 果树学报, 2014, 31(3): 448-453, 340.
葛永红, 李灿婴, 吕静祎, 等. 果蔬采后病原真菌分泌胞外酶的研究进展[J]. 食品科学, 2016, 37(15): 265-270.
胡会刚, 孙光明, 董 晨, 等. 菠萝采后主要病害发生及防治研究进展[J]. 广东农业科学, 2012, 39(24): 93-96.
Wijesinghe C J, Wilson Wijeratnam R S, Samarasekara J K R R, et al. Biological control of Thielaviopsis paradoxa on pineapple by an isolate of Trichoderma asperellum[J]. Biological Control, 2010, 53(3): 285-290.
Wilson Wijeratnam R S, Hewajulige I G N, Abeyratne N. Postharvest hot water treatment for the control of Thielaviopsis black rot of pineapple[J]. Postharvest Biology and Technology, 2005, 36(3): 323-327.
Wijesinghe C J, Wilson Wijeratnam R S, Samarasekara J K R R, et al. Development of a formulation of Trichoderma asperellum to control black rot disease on pineapple caused by (Thielaviopsis paradoxa)[J]. Crop Protection, 2011, 30(3): 300-306.
剛成诚, 李建龙, 王亦佳, 等. 利用不同化学方法处理水蜜桃保鲜效果的对比研究[J]. 食品科学, 2012, 33(6): 269-273.
Conway W S, Sams C E. Calcium infiltration of Golden Delicious apples and its effect on decay[J]. Phytopathology, 1983, 73: 1068-1071.
Conway W S, Greene G M, Hickey K D. Effects of preharvest and postharvest calcium treatments of peaches on decay caused by Monilinia fructicola[J]. Plant Disease, 1987, 71(12): 1084-1086.
Yu T, Yu C, Lu H P, et al. Effect of Cryptococcus laurentii and calcium chloride on control of Penicillium expansum and Botrytis cinerea infections in pear fruit[J]. Biological Control, 2012, 61(2): 169-175.
陈 敏, 高云慨, 宋海超, 等. 氯化钙结合季也蒙毕赤酵母(Meyerozyma guilliermondii)对抑制芒果采后炭疽病效果的影响[J]. 食品科学, 2016, 37(2): 204-209.
Holb I J, Balla B, Vámos A, et al. Influence of preharvest calcium applications, fruit injury, and storage atmospheres on postharvest brown rot of apple[J]. Postharvest Biology and Technology, 2012, 67: 29-36.
Zhao Y, Wang C. Effect of calcium chloride in combination with salicylic acid on post-harvest freshness of apples[J]. Food Science and Biotechnology, 2015, 24(3): 1139-1146.
吴芳芳, 郑有飞, 吴荣军. 钙盐协同枯草芽孢杆菌对苹果采后炭疽病的控制[J]. 植物保护学报, 2009, 36(3): 225-228.
赵 妍, 杨 超, 王若兰, 等. CaCl2处理对苹果采后品质及炭疽病害的影响[J]. 食品与机械, 2013, 29(6): 179-182, 186.
张大智, 詹儒林, 柳 凤, 等. 杧果细菌性角斑病菌细胞壁降解酶的致病作用[J]. 果树学报, 2016, 33(5): 585-593.
吴芳芳, 郑有飞, 万长健, 等. UV-B 辐射增强对苹果采后炭疽病发病情况和抗病相关酶活性的影响[J]. 生态环境, 2008, 17(3): 962-965.
Zhang Z Q, Tian S P, Zhu Z, et al. Effects of 1-methylcyclopropene (1-MCP) on ripening and resistance of jujube (Zizyphus jujuba cv. Huping) fruit against postharvest disease[J]. LWT-Food Science and Technology, 2012, 45(1): 13-19.
杨冬平, 高兆银, 李 敏, 等. 水杨酸结合超声波处理对芒果采后抗病性的影响[J]. 热带作物学报, 2014, 35(5): 974-979.
马桂珍, 高会兰, 张拥华, 等. 链孢粘帚霉几丁质酶的诱导及其抗真菌活性研究[J]. 微生物学通报, 2007, 34(5): 905-908.
李世贵, 顾金刚, 姜瑞波, 等. 生防木霉菌β-1,3-葡聚糖酶活性研究[J]. 微生物学杂志, 2010, 30(4): 88-91.
Chardonnet C O, Sams C E, Trigiano R N, et al. Variability of three isolates of Botrytis cinerea affects the inhibitory effects of calcium on this fungus[J]. Phytopathology, 2000, 90(7): 769-774.
Ortiz A, Graell J, Lara I. Cell wall-modifying enzymes and firmness loss in ripening ‘Golden Reinders apples: A comparison between calcium dips and ULO storage[J]. Food Chemistry, 2011, 128(4): 1072-1079.
张 强, 代文婷, 金新文. 氯化钙与1-甲基环丙烯对伽师瓜果实软化与果胶酶活性及其基因表达影响[J]. 食品与发酵工业, 2019, 45(2): 45-52.
Ranjbar S, Rahemi M, Ramezanian A. Comparison of nano-calcium and calcium chloride spray on postharvest quality and cell wall enzymes activity in apple cv. Red Delicious[J]. Scientia Horticulturae, 2018, 240: 57-64.
毕 阳, 刘红霞. 钙对果蔬采后腐烂的控制及作用[J]. 甘肃农业大学学报, 2000, 35(1): 1-5.
Mansourbahmani S, Ghareyazie B, Kalatejari S, et al. Effect of post-harvest UV-C irradiation and calcium chloride on enzymatic activity and decay of tomato (Lycopersicon esculentum L.) fruit during storage[J]. Journal of Integrative Agriculture, 2017, 16(9): 2093-2100.
Figueroa C R, Opazo M C, Vera P, et al. Effect of postharvest treatment of calcium and auxin on cell wall composition and expression of cell wall-modifying genes in the Chilean strawberry (Fragaria chiloensis) fruit[J]. Food Chemistry, 2012, 132(4): 2014-2022.
李 波, 王 軍, 孙 思. 植物诱导抗病机制的研究进展[J]. 中国植保导刊, 2013, 33(9): 19-24.
向婧姝. 植物过敏性反应分子机制研究进展[J]. 江西农业学报, 2018, 30(12): 41-46.
Obrien J A, Daudi A, Butt V S, et al. Reactive oxygen species and their role in plant defence and cell wall metabolism[J]. Planta, 2012, 236(3): 765-779.
汪开拓, 廖云霞, 袁坤明, 等. β-氨基丁酸处理对桃果实采后灰霉病的影响及其诱导抗病模式研究[J]. 食品与发酵工业, 2016, 42(2): 65-71.
廖云霞, 费良航, 夏明星, 等. 不同浓度β-氨基丁酸处理对葡萄果实抗病性的诱导模式[J]. 食品科学, 2018, 39(17): 221-228.
李会佳, 朱露露, 李 帅, 等. BABA, BTH 对番茄白粉病的抗性研究[J]. 浙江农业学报, 2016, 28(4): 580-585.
