Light Fertilization Affects Growth and Photosynthesis in Mung Bean(Vigna radiata)Plants
2019-01-09ChiaraAmitranoErmenegildaVitaleVeronicaDeMiccoCarmenArena
Chiara Amitrano,Ermenegilda Vitale,Veronica De Micco,Carmen Arena†
1Department of Agricultural Sciences,University of Naples Federico II,Via Università 100,Portici(NA),Italy
2Department of Biology,University of Naples Federico II,80125 Naples,Italy
Keywords Mung bean Light quality Photosynthesis Crop production Proteins
Abstract Inaclimatechangescenario,theoptimizationofgrowthconditionsforfood crop species plays a key role for the sustainability of cultivation.Agrotechnologies need to be improved to set up the best conditions to maximize plant development,production and resource use efficiency in growth chambers and greenhouses.The manipulation of light quality during plant growth may be used as a powerful mean to obtain specific functional traits.This practice may be useful to improve plant growth,also avoiding the use of large doses of chemical fertilizers,which may compromise the environment and human health.In our study,we analyzed specific physiological traits of mung bean(Vigna radiata)seed-lings grown at different light quality regimes(W-White,R-Red and RB-Red-Blue light),to assess the best light treatment in promoting plant development and photosynthesis.Plant growth was monitored measuring stem and root elongation,dry biomass and total leaf area.The integrity of the photosynthetic machinery was monitored through fluorescence an emission measurements and content of photosynthetic pigments and total proteins.Our results showed that the growth under R wavelengths promoted stem elongation comparedto W andRB.Thislighttreatmentwas alsoresponsibleforthehighestproduction of total chlorophylls.Photochemistrywas not affectedby the differentlight qualities.RB light induced a compact architecture of plants and the highest amount of proteins.Overall results indicate that different light quality regimes can be applied during the cultivation to consciously modify plant growth and development.Thus,it will be fundamental to optimize and choose opportunelynot only the intensity but also the spectral composition of light to maximize the productivity of a specific crop in quantitative and qualitative terms.
1 Introduction
The optimization of production processes infood crops is afundamental issue in the perspective ofthe increasing demand in quantity and quality of food(Lairon,2010).
Vigna radiata(L.)R.Wilczek also known as mung bean,is an important legume crop in a large number of countries(Lakhanpaul et al.,2000).The seeds and sprouts of mung bean have been consumed as common food in China for more than 2000 years;they are used in fresh salads in India,Bangladesh,South East Asia and western countries providing the main source of protein(240 g kg-1)for a large proportion of vegetarians as well as abundant dietary fiber,carbohydrates(630 g kg-1)and a vast range of bioactive phytochemicals with antioxidant,antimicrobial,anti-in flammatory and antitumor activity(Kanatt et al.,2011;Nair et al.,2013;Tang et al.,2014).Mung bean can be considered an excellent example of functional food because it presents a high digestibility and it is rich in proteins(about 20%-24%of the total composition of mung bean)and essential amino acids(Kudre et al.,2013;Tang et al.,2014).In this species,seeds germination and formation of sprouts are processes strongly in fluenced by light that represents the main driver for plant development, flower and seed production(Singh et al.,2015).In fact,it is well known that some useful metabolites are synthesized during the initial stage of germination(El-Adawy et al.,2003).Beside germination,light quality and intensity affect numerous other processes,such as photo-morphogenesis,biomass accumulation,photosynthetic efficiency, flowering and phytochemical synthesis.In particular light quality and intensity modify the signaling cascade of specific photoreceptors(phytochromes,cryptochromes and phytotropins),thus changing the expression of a high number of genes(Yorio et al.,2001;Massa et al.,2008;Li and Kubota,2009;Lin et al.,2013;Olle and Viršile 2013;Arena et al.,2016).Within the whole visible light spectrum,pure red wavelengths strongly affect the vegetative growth,photosynthetic apparatus development,photosynthetic process,morphogenesis, flowering and budding;pure blue wavelengths play a fundamental role in the regulation of vegetative growth and photosynthesis through the control of chlorophyll biosynthesis,stomata opening and photo-morphogenesis(Urbonavičiute,et al.,2007;Chen et al.,2014;Singh et al.,2015).Plants have a considerable physiological and morphological plasticity in adapting to different light qualities(Barreiro et al.,1992;Arena et al.,2016).In particular blue light combined with red wavelengths positively in fluences plant growth,tissue nutritional value,chlorophyll ratio and number of seeds in vegetables(Yanagi et al.,1996;Goins et al.,1997;Li et al.,2012).It is not possible to delineate major common responses for all crops because there are species-specific reactions to light quality and intensity(Li,2012;Singh et al.,2015).
The modulation of light quality is more and more recognized as an efficient mean to optimize not only plant development but also nutritional quality of edible organs.At present,many greenhouses are equipped with arti ficial light provided by LED technology,which in combination with solar light as the main light source guarantee an optimal illumination during plant growth(Bian et al.,2015).A proper lighting during plant development is a valuable resource,especially in regions where solar radiation is weak or photoperiod is not sufficient to guarantee optimal growth(Opdam et al.,2005).Cultivation under controlled conditions plays an important role in the production of vegetables:the main advantage provided by the indoor production is the control of the growth environment in terms of light(quality and quantity),temperature,humidity,CO2concentration,water and nutrients(Gary,2003;Gruda,2005).For light optimization,the LEDs technology used in artificial systems is a suitable device easily integrated into digital control systems to modulate both the intensity or spectral composition during the whole plant life cycle(Yeh and Chung,2009).The manipulation of LED lighting represents a useful mean to realize appropriate light fertilization protocols in order to satisfy specific light requirements for each species.This is a fundamental requisite to obtain high photosynthetic rates and biomass production without the addition of high doses of chemicals.An appropriate light fertilization is “eco-friendly”because allows minimizing the use of fertilizers to obtain high crop yield,thus reducing the impact on the environment.For example,the modulation of light quality regulates the nitrate concentration in plants:red light stimulates the nitrate reductase activity decreasing the nitrate concentration in plants,whereas blue light increases the overall nitrogen concentration,reducing the plant demand for this macronutrient(Deng et al.,2000;Lillo and Appenroth,2001;Ohashi-Kaneko et al.,2006).The selection of a proper light spectrum in growth chambers or in greenhouses in combination with the most suitable environmental parameters(T◦,RH%,CO2concentration)is promising to achieve high yield in a short time.In this study we applied specific “light-fertilization”treatments to improve the growth and quality of mung bean(Vigna radiata L.)seedlings.To this purpose,we analyzed plant development,photosynthesis,biomass partitioning,chlorophyll and total protein production,providing useful information for the design of light systems for the cultivation under controlled conditions.
2 Materials and methods
2.1 Plant material and experimental design
Seeds of Vigna radiata(L.)R.Wilczek,commonly known as mung bean,were incubated in Petri dishes,lined with three layers of filter paper,at 22◦C in the dark for two days.Afterward,germinated seeds were incubated in a growth chamber under controlled conditions of temperature(22-23◦C),relative humidity(60-65%)and photoperiod(12 h),under three different regimes of light,equivalent for intensity(photosynthetic photon flux density,PPFD,140-200µmol photons m-2s-1),but different for quality:white light(W)with a spectrum from 380-750 nm,pure red light(R)with a maximum intensity at 660±5 nm,and a mixture of red(R:66%)and blue(B:33%)light(RB).The white light was provided by two types of Phosphor-Coated LEDs for white light(5000K-6500K).
2.2 Plant growth
After a week,at the emergence of the first two true leaves,10 seedlings for each light treatment were transplanted into pots filled with commercial soil and incubated for four weeks,under the three conditions reported above.During the growth period,seedlings were regularly watered to reintegrate water lost by evapotranspiration.Plant height,stem and root length,root/shoot ratio,leaf number,leaf area and plant dry weight were recorded at the end of the experiment.The leaf area(surface of the lamina)was quantified on digital images taken by means of a digital camera and analyzed through “ImageJ”software(Rasband,W.S.,U.S.NIH,Bethesda,Maryland,USA,1997-2012).
2.3 Fluorescence“a” emission
Chlorophyll a fluorescence emission was measured using a pulse amplitude modulated fluorometer(Junior-PAM,Walz,Germany).Light fast-kinetic response curves(LFRCs)were performed in mung bean seedlings exposed to different light treatments in order to evaluate the photosynthetic capacity at Photochemical Photon FluxDensities(PPFD)ranging from 0to1500µmolephotons m-2s-1.Foreach irradiance level,photochemical parameters,in the dark and in the light,were calculated as reported in Arena et al.(2005).
On 30 min dark-adapted leaves,the background fluorescence signal,F0,was induced by a weak light of 0.5µmol photons m-2s-1at 0.6 kHz frequency.Maximal fluorescence in the dark-adapted state,Fm,was obtained by applying a 1 s saturating light pulse(8,000µmol photons m-2s-1)at 20 kHz frequency.The photochemical quenching qP,which indicates the proportion on PSII reaction centers that are open,was calculated according to Genty et al.(1989).
The non-photochemical quenching(qN),the fraction of absorbed light energy not utilized in photochemistry and dissipated as heat was calculated according to Van Kooten and Snel(1990).
2.4 Content of chlorophylls and proteins
After fluorescence measurements,5 leaves were collected from 3 plants per each treatment for the determination of photosynthetic pigment content.The content of chlorophylls a and b,the relative a/b ratio,and total carotenoids(x+c)were determined.Photosynthetic pigments were extracted in ice-cold 100%acetone,quantified and expressed inµg cm-2as reported in Arena et al.(2014).
For protein extraction,0.3 g of leaves from seedlings per each light treatment were powdered with liquid nitrogen.Proteins from the cytoplasmic fraction,were extracted from the powder according to Wang et al.(2006)and Sorrentino et al.(2018).Proteins were estimated by the method of Bradford(1976)based on the colorimetric assay with Coomassie blue R-250,using the Bio-Rad Protein Assay(Bio-Rad).A standard curve with bovine serum albumin(Sigma)was used to calculate protein concentrations in the extracts,expressed in mol[BSA]g-1FW.
2.5 Data elaboration
All data were analyzed by one-way analysis of variance(ANOVA)through the Sigma-Stat 3.5 software(Jandel Scientific,San Rafael,CA,USA).The Holm-Sidak test was applied for all multiple comparison tests with a significance level of p<0.05.Data are reported as mean values±standard error(n=5).
3 Results and discussion
3.1 Effect of light quality on plant growth
V.radiata seedlings grown under R light showed a significantly higher values(P<0.05)of plant height and leaf area than those grown under W and RB regimes(Fig.1a,e).These results are in agreement with other studies where the stem elongation in Chrysanthemum(Kim et al.,2004c)and grape(Puspa et al.,2008)was stimulated by red light.The addition of blue to red wavelengths determined significant reduction(P<0.05)in stem elongation and in root/shoot ratio,accompanied by an increase in leaf number compared to the other light treatments(Fig.1a,c,d).The high root/shoot ratio is due to the significant increase in root length recorded in RB seedlings compared to W and R ones(Fig.1b).These results are in agreement with previous studies which reported an increment in shoot length under 100%of red light(Hoenecke et al.,1992;Miyashita et al.,1997;Brown et al.,1995;Goins et al.,1997;Okamoto et al.,1997).Although both red and blue light can mediate stem elongation(Huche-Thelier et al.,2016),many studies have shown that blue light was more effective than red light in suppressing shoot elongation in a range of plant species(Hoenecke et al.,1992;Brown et al.,1995;Kong et al.,2012).
Generally,different plant species show different responses to specific light receipts(Goins et al.,1997):for some crops,such as lettuce(Lactuca sativa L.),spinach(Spinacia oleracea L.)and radish(Raphanus sativas L.),the growth under pure Red LEDs was not sufficient to achieve a normal development root/shoot ratio and a minimum blue light was necessary to guarantee this balancing(Bula et al.,1991;Yorio et al.,2001).
The growth under RB wavelengths induced a more compact structure characterized by the short stem elongation and the higher number of smaller leaves(at least compared to the R seedlings).In these plants,the blue light component added to the red one likely promoted the partitioning of photosynthates to roots,conversely to R plants where the total absence of blue determined the unbalancing of photosynthetic products towards the shoot.The increasing in root length under RB light and the rise of stem elongation under R light were responsible of the higher values(P<0.05)of total dry weight in RB and R seedlings compared to W seedlings(Fig.1f).
The higher partitioning of photosynthates towards the epigeal biomass in R compared to W plants as well as the increased leaf area expansion may be likely due to the need by photosynthetic apparatus to harvest as much light as possible,since the pure R light does not represent a favorable condition for plant growth.When blue light is added to the red wavelengths,the stomatal opening is promoted and thus gas exchanges are improved,resulting in increased production of biomass in many species(Sharkey and Raschke,1981;Goins et al.,1997;Zeiger and Zhu,1998).
Okamoto et al.(1997)reported that the stem elongation in lettuce seedlings decreases significantly with the increase of blue wavelengths.It is well demonstrated that small variation within blue light component can determine significant changes in stem growth(Brown et al.,1995).The synergistic interaction of the blue and red lights receptors(cryptochromes and phytochromes)may account for such responses(Kim et al.,2004 a,b,c).Also in our experiment,even if the light intensity was the same during plant growth for all light treatments,the different light quality may have stimulated specific responses,mediated by diverse photoreceptors,that make seedling able to optimize their growth according to the specific light environment.
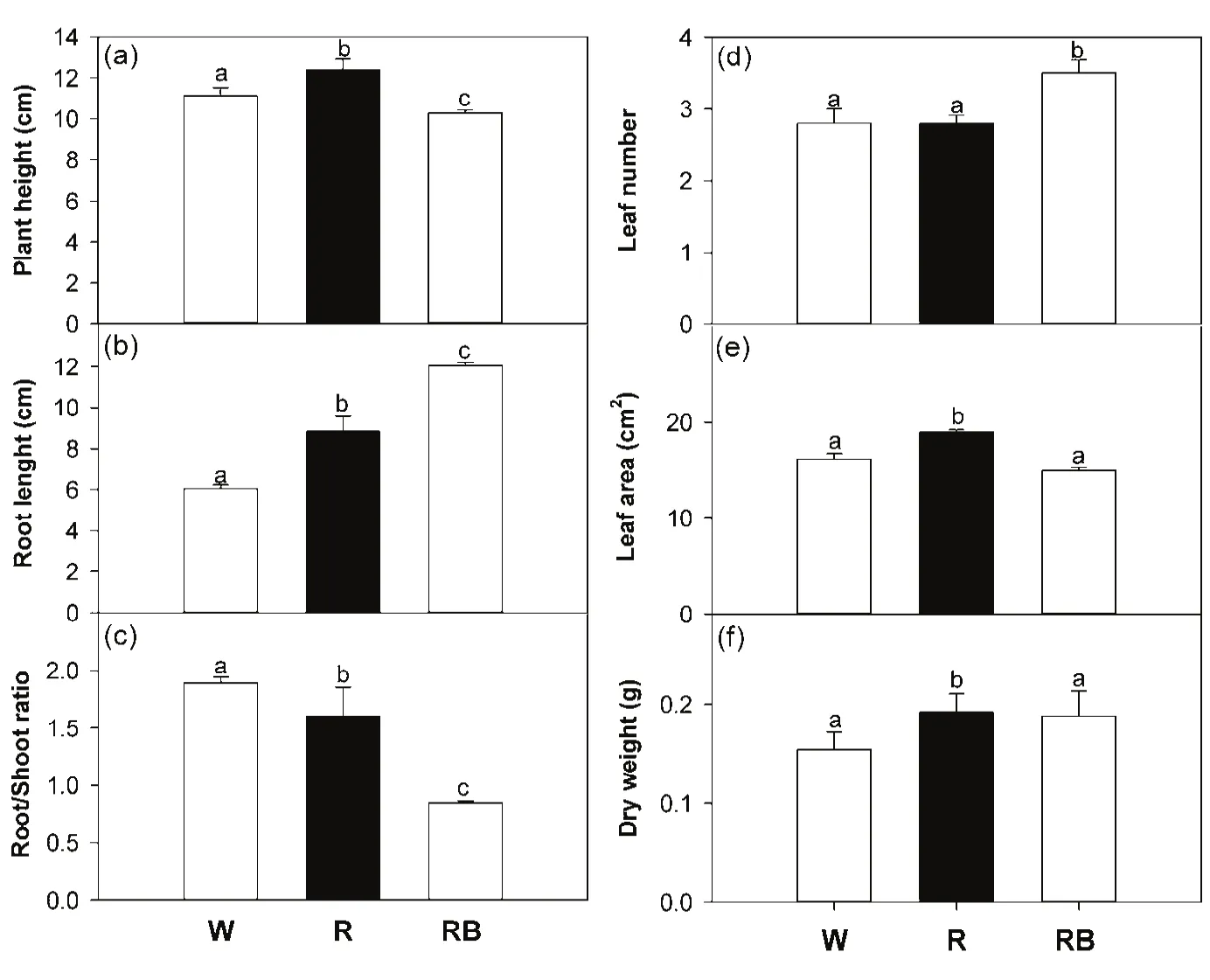
Fig.1 Plant height(a),root length(b),root/shoot ratio(c),leaf number(d),leaf area(e),and dry weight(f)of Vigna radiata L.seedlings grown under the three different light quality treatments:White(W),Red(R)and Red-Blue(RB).Each value represents the mean±SE(n=5).Different letters indicate significant differences between light treatments(P<0.05).
3.2 Effect of light quality on photochemistry
The photosynthetic efficiency of seedlings subjected to the different light-growth regimes was analyzed in vivo by fluorescence emission measurements.Light fast-kinetic response curves have been performed to assess if the capability of photosynthetic apparatus in converting the light energy to photosystems may be affected by light quality.Ft showed an increase according to the increasing irradiation in all seedlings,irrespective of the specific light quality treatment.However,in W seedlings the Ft values were significantly higher(P<0.01)than in R and RB seedlings not only at growth irradiance of 200µmol photons m-2s-1(see the bar plots inside the graph)but in the whole range of PPFD tested(Fig.2a).On the other hand,the photochemical quenching,qP,exhibited a progressive reduction as irradiation increased.At growth irradiance,qP was higher(P<0.05)in R and RB than in W seedlings(Fig.2b).The decline in photochemical quenching with photon flux density increase was accompanied by the rise of non-photochemical quenching(qN)(Fig.3c).RB seedlings showed the highest values(P<0.01)of qN compared to W and R in the whole range of PPFD,including the plant growth irradiance(Fig.2c).The progressive increase of Ft together with lower qP indicated a loss of photosynthetic efficiency in W compared to R and RB seedlings.It has been demonstrated that the reduction of photochemical reactions leads to the depletion of PSII reaction center function(Maxwell et al.,1994;Wang et al.,2009),resulting in decline of plant productivity.
Generally,the decrease in photochemistry is balanced by the rise of non-photochemical quenching that acts as compensation mechanism to avoid an over excitation of photosynthetic electron transport chain(Maxwell and Johnson 2000).This is not the case or V.radiata plants exposed to white light,in which the lowest values of qN indicates a reduced capacity to dissipate thermally the excess of light energy to photosystems.On the other hand,the opposite common behavior of R and RB seedlings(higher qP values)confirms the positive role of R and B wavelengths on light reactions of photosynthesis.In previous studies,highest values of photochemical quenching were found in tomato seedlings grown under RB condition compared to white and pure red light(Liu et al.,2011).Conversely to other species,such as Cucumis sativus and Acacia mangium(Wang et al.,2009;Yu and Ong,2003),the growth under pure red did not induce a decline of qP compared to white light in V.radiata seedlings,highlighting the species-specific response to light quality.
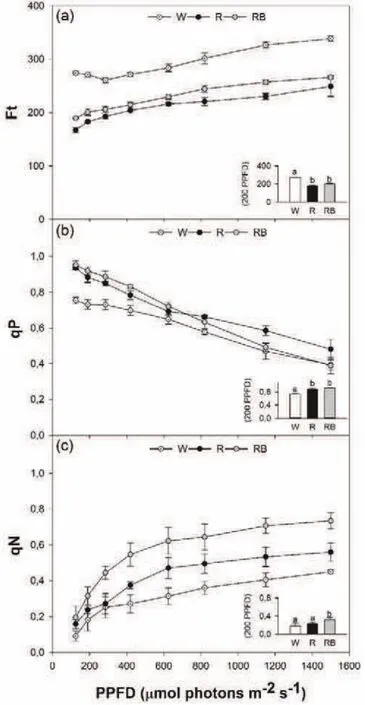
Fig.2 Fast kinetics light-response curves(from 125 to 1500µmol photons m-2s-1)of Ft-basal fluorescence in the light(a),qP-photochemical quenching(b),qN-non-photochemical quenching(c)of Vigna radiata L.seedlings grown under three different light quality regimens:white(W),Red(R)and Red-Blue(RB).The bar plot diagram enclose in each fast kinetics light-response curve represents the value of the parameter at growth irradiance in chamber of 200µmol photons m-2s-1.Each point is the mean±SE(n=5).Different letters indicate significant differences among light treatments(P<0.05).
In the specific case of V.radiata seedlings,the growth under RB wavelengths stimulates the thermal dissipation mechanisms not reducing the photochemical reactions.This could be a favorable trait because it makes RB seedlings stronger against photoinhibitory damage risks.It is well known that an increase of qN may occur as photoprotective process against light-induced damages(Hong et al.,1999;Maxwell and Johnson,2000).
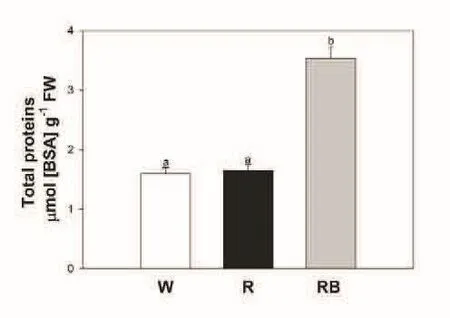
Fig.3 Total protein content in Vigna radiata L.plants grown under the three different light quality treatments:White(W),Red(R)and Red-Blue(RB).Each value represents the mean±SE(n=5)and is referred to a BSA(albumin serum bovine)standard curve,at the wavelength of 595nm.Different letters indicate significantly different values(P<0.05).
3.3 Effect of light quality on pigments and total proteins
The different light quality treatments during growth significantly affected the content of photosynthetic pigments in V.radiata seedlings(Table 1).Compared to W and R,RB seedlings showed lower values(P<0.05)of chlorophyll a and b and total carotenoid content as well as Chl a/b ratio.Compared to W,seedlings developed under pure R exhibited similar values for chlorophyll content and chlorophyll a/b ratio and higher(P<0.01)total carotenoid(x+c)content(Table 1).The higher amount of chlorophylls and carotenoids in R than RB plants together with the higher values of leaf area and stem elongation,compared to W seedlings,may be interpreted as a strategy to optimize the light harvesting by photosynthetic apparatus under limiting light conditions.
Our hypothesis is in agreement with data reporting that an increase in photosynthetic pigment synthesis is often associated with an increase in leaf lamina expansion to improve light interception(Kubota et al.,2012).In W plants the high pigment content was not accompanied by an increase in stem elongation and leaf area,probably because the mix of different wavelengths is more suitable to guarantee the right amount of light to photosystems reducing the need for allocating biomass in above-ground organs.The lowest(P<0.05)chlorophyll a/b ratio in RB seedlings is indicator of a different acclimation to light in these plants when the red wavelengths are supplemented with blue.This parameter may be considered as a“bioassay for the light environment of a plant”(Dale and Causton,1992).In our experiment the significant reduction of the chlorophyll a/b ratio under RB treatment compared to W and R,indicating a simultaneous decrease of both chlorophyll a and b;this may represent a signal that the light at leaf lamina is not limiting for photochemical reactions in RB seedlings.
Regarding the protein content,results from the Bradford assay indicated that the highest amount was found in RB seedlings which showed significantly higher values(p<0.05)than R and W seedlings(Fig.3).This is in agreement with previous experiments showing that the blue wavelengths promote protein synthesis and accumulation(Zhang et al.,2010;Li et al.,2012).The stimulatory effect on proteins played by RB conditions makes this light regime interesting to induce a valuable trait in V.radiate seedlings.
4 Conclusions
The different light quality significantly in fluenced specific traits in V.radiata seedlings.The increasing in root growth under RB light,and the rise in stem elongation under R light,lead to higher values of total dry weight in RB and R seedlings compared to W seedlings.Moreover,the development of plants under the mix of RB lights,induced a more compact structure,very helpful in the sight of its cultivation in growth chambers when volume availability is a constrain.
When blue wavelengths are mixed with R ones,not only the capability of photosynthetic apparatus to use the light energy is enhanced but also the thermal dissipation mechanisms are promoted in V.radiata.From the eco-physiological point of view,this trait is important to give these plants an intrinsic resistance against photoinhibition risks.Finally,compared to W and R treatments,RB light regime enhances the content of total proteins.This outcome could be very important from a nutraceutical point of view.However,further analyses are needed,also considering light quality effects on edible organs also in other species.
The overall results indicate that it is possible to use specific light fertilization treatments as a mean to drive crop growth and development,thus obtaining desirable traits in plants.The application of different light spectral composition may complement and/or reduce the utilization of other growth-promoting factors.However,these outcomes seem to be highly depending on the species and this aspect has to be adequately considered.
杂志排行

Journal of Environmental Accounting and Management的其它文章
- Capture Rate of Selected Heavy Metals In Q.Ilex L.Leaves Collected At Two Sites With Different Land Uses
- Impact of Biochar Amendment on Soil Quality and Crop Yield in a Greenhouse Environment
- Regional Redistribution Effects of Renewable Energy Subsidies
- Phytotoxic Extracts as Possible Additive in Subsurface Irrigation Drip for Organic Agriculture
- Ecotoxicological Assessment of Virgin Plastic Pellet Leachates in Freshwater Matrices
- Uptake of Micro and Macronutrients in Relation to Increasing Mn Concentrations in Cistus salvifolius L.Grown in Hydroponic Cultures