棉花WRKY22基因分离及其对黄萎病的抗性分析
2019-01-09张振楠刘建芬司怀军吴家和
雷 煜,张振楠,胡 广,刘建芬,唐 叶,张 宁,司怀军,吴家和
(1.甘肃农业大学 生命科学技术学院,甘肃 兰州 730070;2.中国科学院 微生物研究所,北京 100101; 3.棉花生物学国家重点实验室,郑州大学基地,河南 郑州 450001)
植物不能够像动物那样趋利避害的自由移动,因此,当病原菌浸染时,其只能通过调节自身的生理生化反应来抵抗或适应这些变化,这些变化被称为植物先天免疫反应。一般认为植物的先天免疫反应包括2个层次或进程,一个是由病原菌相关分子模式(PAMP)启动的先天免疫反应,称为PTI,另一个是由病原菌效应因子(Effector)启动的先天免疫反应,称之为ETI[1-2]。当植物先天免疫反应时细胞内的抗病相关基因表达发生了改变(Reprogram),而这些抗病相关基因主要是受编码调控蛋白和转录因子调控的,其中转录因子主要包括WKRY、MYB、NAC、bZIP等[3]。
WRKY转录因子是植物中最大转录因子家族之一,在拟南芥中有72个WRKY转录因子[4],棉花中报道的为116个[5]。WRKY蛋白一般被分为3个组群,分别为GroupⅠ、Ⅱ、Ⅲ,其中Group Ⅱ又被分为5个亚组,被标为Ⅱa、b、c、d和e。WRKY蛋白含有WRKYGQK保守结构域,其C端含有C2H2或C2C2的锌指结构[5-6]。WRKY转录因子通常作为植物信号网络中的重要组成因子,在植物体内承担着多种功能,包括植株的生长发育和逆境响应。其中对病原菌抗性反应研究涉及多个WRKY转录因子,如在拟南芥中,WRKY7、33、3、22、48、70、72和40等直接参与抗病反应[7-14],这些蛋白有的参与腐生性病原菌抗性,有的参与寄生性病原菌的抗性,有的对二者都能产生抗性反应[15]。这些WRKY转录因子的抗病机制比较复杂,不断增加的研究报道说明,WRKY转录因子可以通过多种途径接受抗病信号,从而调控下游抗病靶基因的表达。
棉花是一种重要的经济作物,属于人们种植的主要作物之一[16]。棉花纤维是纺织工业的重要原料,是支撑纺织工业发展和人们生活的基础;同时棉籽油是食品和化工用油的主要来源之一,棉籽粕是一种牛羊等反刍动物的蛋白饲料添加成分[17]。棉花稳产性是保障种植效益的首要因素,然而,由大丽轮枝菌(Verticilliumdahliae)引起的棉花黄萎病是影响当前棉花稳产性的最重要因素之一,因此,如何提高棉花品种的抗病性是当今棉花育种的一个主要目标。有关棉花抗病WRKY转录因子的研究已经有一些报道,如GhWRKY3[18]、GhWRKY40[19]和 GhWRKY1[20]。然而,棉花有116个WRKY蛋白,参与棉花抗病反应的WRKY蛋白鉴定和其抗病反应的分子机制等有待于进一步研究。
本研究中,通过比对拟南芥中抗病相关的WRKY蛋白序列,分别在陆地棉中获得这些蛋白的直系同源基因,分析这些GhWRKY基因对大丽轮枝菌侵染时的表达响应。筛选出了对病原菌反应剧烈的GhWRKY22基因,并对其进行基因沉默(Knockdown)的研究。结果显示,GhWRKY22基因沉默的植株对大丽轮枝菌的抗性下降,同时抗病标记基因(Marker genes)PDF1.2、PR1、PR3、PR4、PR5和PAL等的表达分析结果表明,GhWRKY22通过水杨酸(SA)和茉莉酸(JA)信号途径参与大丽轮枝菌抗性反应。总之,GhWRKY22基因正调控棉花对黄萎病的抗性,可以作为棉花抗病候选基因用于棉花的抗病育种。
1 材料和方法
1.1 试验材料
陆地棉遗传标准系TM-1(Gossypiumhirsutumacc. TM-1)以及棉花抗黄萎病品系BD18由山西省农业科学院棉花研究所罗晓丽研究员提供。
根癌农杆菌菌株 GV3101、大肠杆菌菌株 DH5α,均由中国科学院微生物研究所植物基因组国家重点实验室保存。
强致病力落叶型黄萎病菌菌株 V991 (Verticilliumdahliaestrain,V991),由中国农业科学院植物保护研究所简桂良研究员惠赠。
植物病毒诱导基因沉默(Virus induced gene silencing,VIGS)技术系统,包括干扰表达载体 pYL156以及辅助载体 pYL192由清华大学刘玉乐教授馈赠。
1.2 基因序列比对分析与进化树构建
根据文献报道从模式植物拟南芥中选取抗病相关的WRKY编码蛋白作为对象,在棉花数据库(https://www.cottongen.org)中进行Blast同源比对,获得了8个棉花直系同源WRKY编码蛋白,利用Clustalx 2.1软件进行氨基酸多重比对,并使用 MEGA 5.0邻接法生成系统进化树进行同源性分析。
1.3 棉花材料的培养
将干燥的硫酸脱绒种子在水中37 ℃浸泡过夜,移至培养盒中并在其上覆盖2~3 cm厚的灭菌湿润的蛭石,置于暗处,每天补充适量水分以保证种子正常萌发。大约3 d后,将长势一致的棉花幼苗移植到注满水的12孔水培盒中,放置于16 h光照/8 h黑暗、25 ℃、光强120 μmol/(m2·s)的培养箱中进行培养。每天定时观察适当加水确保棉苗正常生长,以备接菌和取样。
1.4 棉花根部RNA的提取及cDNA合成
剪取不同处理棉花样品,立即置于含液氮的研钵中迅速研磨成粉状,随后使用植物总RNA分离试剂盒(Sangon,上海)提取和纯化组织样品的总RNA,并用Nanodrop检测RNA样品的纯度与浓度,其后通过1.0% 的琼脂糖凝胶电泳检测总RNA 的完整性,选择纯度和完整性优良的RNA进行cDNA合成。按照试剂盒提供的方法,用EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix(Transgen Biotech,北京,中国)将1 μg RNA进行逆转录合成cDNA。反转录获得的cDNA稀释10倍置于-20 ℃冰箱保存。
1.5 基因片段克隆及载体构建
选取GhWRKY22基因特异的核苷酸序列设计引物,并分别在正向引物和反向引物前添加限制性内切酶BamHⅠ和KpnⅠ识别位点以及保护碱基(表1)。用TM-1遗传标准系陆地棉的cDNA为模板进行PCR扩增GhWRKY22基因,利用双酶切法将目的片段插入到植物病毒表达载体 pYL-156中;酶切和测序鉴定正确后命名为pYL-156-GW22。
1.6 农杆菌介导的VIGS植株培育
通过电击法分别将pYL-156-GW22、pYL-156-PDS(阳性对照载体)、pYL-156(空载体)和pYL-192辅助载体转化到农杆菌菌株GV3101中,PCR验证无误备用。
将上述农杆菌菌株在含有50 mg/mL卡那霉素、25 mg/mL利福平的液体LB培养基中于28 ℃,200 r/min培养过夜。菌液在室温下4 000 r/min离心7 min(Eppendorf 5810R,德国)收集农杆菌细胞。随后将沉淀的菌体重新悬浮于MMA(10 mmol/L MES,10 mmol/L MgCl2,200 mmol/L AS)溶液中,调节至终浓度OD600为1.0~1.2,将含有pYL-156-PDS、pYL-156-GW22和pYL-156(空载体)的农杆菌悬浮液分别与pYL-192等比例混合均匀,随后室温暗处孵育3 h备用。当BD18棉花2片子叶完全展开时,选取长势健壮一致的棉花幼苗进行农杆菌注射浸染。先用针头在子叶背面划出伤口,再用1 mL无针注射器将菌液从子叶背部伤口处注入,叶面浸染面积达90%以上为佳。侵染后的棉株室温避光12 h左右,随后移回光照培养箱中进行培养。
1.7 大丽轮枝菌活化和棉花接菌处理及其抗病性调查
将保存于-80 ℃的大丽轮枝菌菌株取适量均匀涂布于PDA培养基上,28 ℃暗培养3 d后,挑取菌块于100 mL查式培养液中,放置在28 ℃ 200 r/min摇床上培养,4~5 d后取出用8层医用纱布过滤菌丝,利用血球计数板计算孢子浓度,并用5%的蔗糖溶液调整其浓度为106个/mL备用。
TM-1棉花接种大丽轮枝菌的方法参考文献[21]报道,简而言之,取长势一致并具有3片真叶的棉株,对其主根进行统一伤根处理,将棉花根部浸泡到上述准备好的菌液中,50 min后放回水培盒中并置于光照培养箱中培养。收取0,12,24,36,48,72,96 h等不同处理时间的根样品进行棉花WRKY基因表达分析。
为了观察棉花植株发病情况,根据范强等[22]报道,采用土培法对基因沉默BD18植株进行接菌,并在接菌21 d后,统计植株的发病棵数和发病程度,计算发病率和病指。参照Wang等[21]报道,进行大丽轮枝菌回复培养试验,简而言之:分别剪取对照组与试验组第一节间的茎段进行表面灭菌(浸泡在25%~50%的84消毒液中5 min)后横向切成小段,放置在PDA培养基上,25 ℃培养5 d后观察这些切断的大丽轮枝菌生长状况。
1.8 SA和JA处理
棉花幼苗具有2片真叶时,分别用茉莉酸甲酯(Methyl jasmonate,MeJA)和水杨酸(Salicylic acid,SA)进行激素诱导处理,具体方法参照先前报道的方法[23]。在激素处理0,0.5,1.0,3.0,6.0,12.0 h后分别取其根系,进行GhWRKY22基因诱导表达分析。
1.9 实时荧光定量PCR(qRT-PCR)分析
以不同棉花样本cDNA为模板和目的基因特异引物(表1)进行qRT-PCR分析,具体反应体系按照TransStart Top Green qPCR SuperMix试剂盒使用说明(Transgen Biotech Beijing,China)进行配置,以棉花持家基因UBQ7为内参基因。反应使用的是IQ5多色检测系统(Bio-Rad,Hercules,CA,USA),反应条件为 95 ℃ 5 min;95 ℃ 30 s,60 ℃ 20 s,72 ℃ 30 s,40个循环。数据处理采用2-ΔΔCt方法计算基因的相对表达量[24],试验重复3次。

表1 引物序列Tab.1 Primer sequence
2 结果与分析
2.1 生物信息学分析筛查棉花抗病相关WRKY基因
WRKY转录因子参与植物生长发育和抗性反应等一系列过程,能够调控多个生理途径基因表达。在拟南芥中,已经报道多个WRKY转录因子参与植株的抗病反应,主要包括WRKY7、33、3、22、48、70、72和40等(表2)。根据8个拟南芥WRKY抗病蛋白序列进行(直系)同源比对,在棉花基因组数据库中筛查到11个棉花WRKY转录因子,如图1所示,其中8个棉花WRKY转录因子分别与拟南芥的8个WRKY响应的聚合在一起,暗示它们可能是直系同源基因。这些抗病相关的WRKY转录因子分布在3个WRKY类型中,在Ⅱ型的5个亚类a、b、c、d和e中也分别存在,说明参与调控抗病反应的WRKY蛋白类型比较广泛。

表2 拟南芥抗病相关WRKY转录因子和其对应棉花直系同源基因Tab.2 Defense related WRKY transcription factors in Arabidopsis and their orthologous genes in cotton
2.2 棉花抗病相关WRKY基因在大丽轮枝菌侵染下的表达响应
棉花黄萎病是棉花的主要病害,其病原菌为大丽轮枝菌,因此,这8个与拟南芥直系同源的棉花WRKY基因表达是否对大丽轮枝菌产生应答响应,是研究这些GhWRKY基因抗病功能的前提。在水培盒中培养21 d大小的棉花苗被用于接菌试验。接种大丽轮枝菌后0,12,24,36,48,72,96 h进行根部取样,通过qRT-PCR方法分析这8个GhWRKY基因的表达情况。相对于0 h对照,其中有6个GhWRKY基因的表达在72 h之前均受到大丽轮枝菌诱导上调,另外2个WRKY基因(GhWRKY48和GhWRKY70)表达变化比较复杂,GhWRKY48的表达量总体呈现下降趋势,而GhWRKY70表现为升-降-升的趋势。在图2中GhWRKY7、GhWRKY40、GhWRKY22、GhWRKY3和GhWRKY72等5个WRKY基因表达量在24 h左右和72 h分别表现为上调,呈现出双峰曲线,暗示这5个WRKY基因的表达量可能受到水杨酸(SA)和茉莉酸(JA)2种激素信号的调控[25]。而GhWRKY48在病原菌浸染下总的表现为下降趋势,说明这个GhWRKY基因是一个负调控抗病的转录因子。
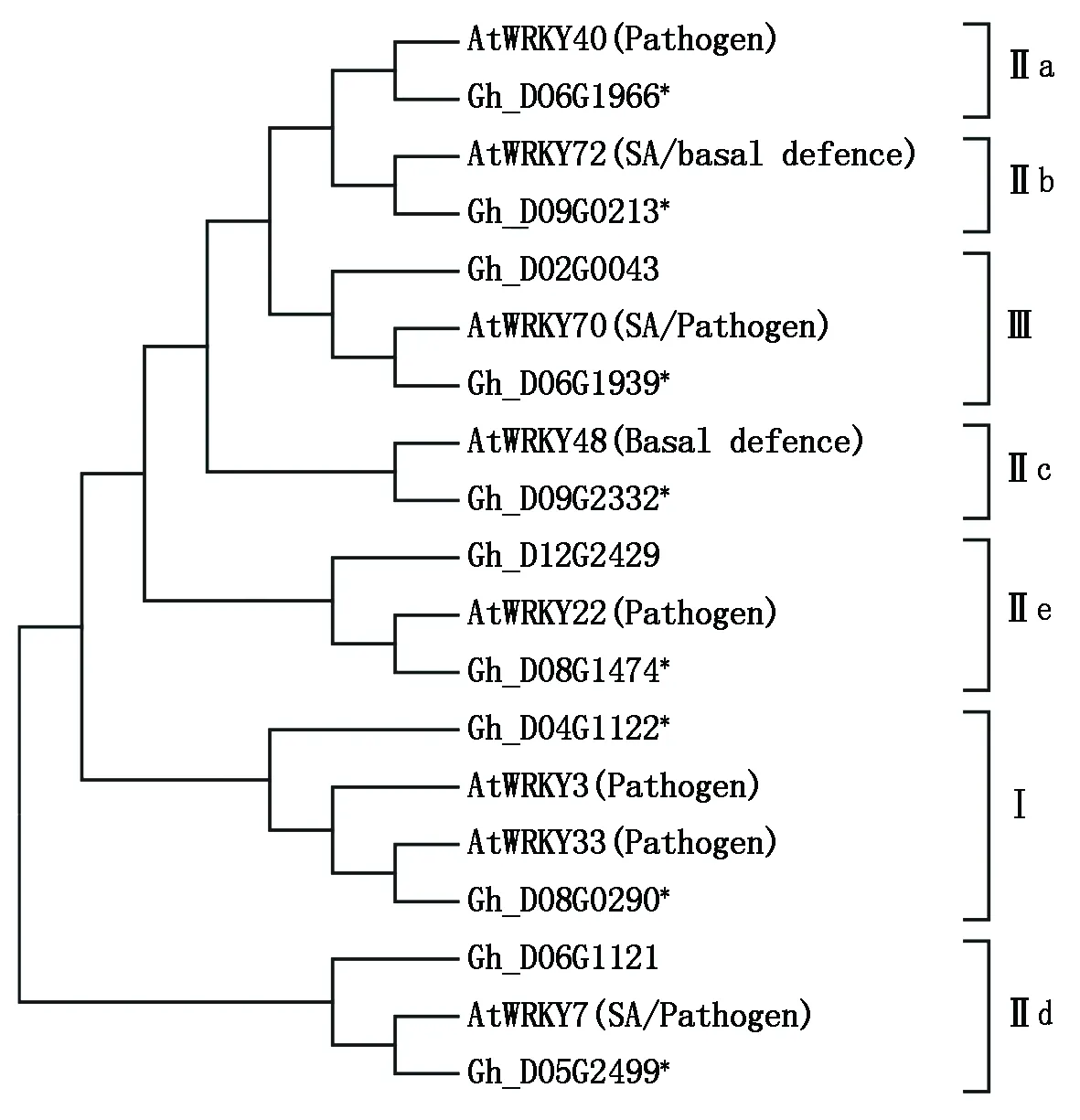
*.棉花直系同源基因。*.Orthologous cotton WRKYs.
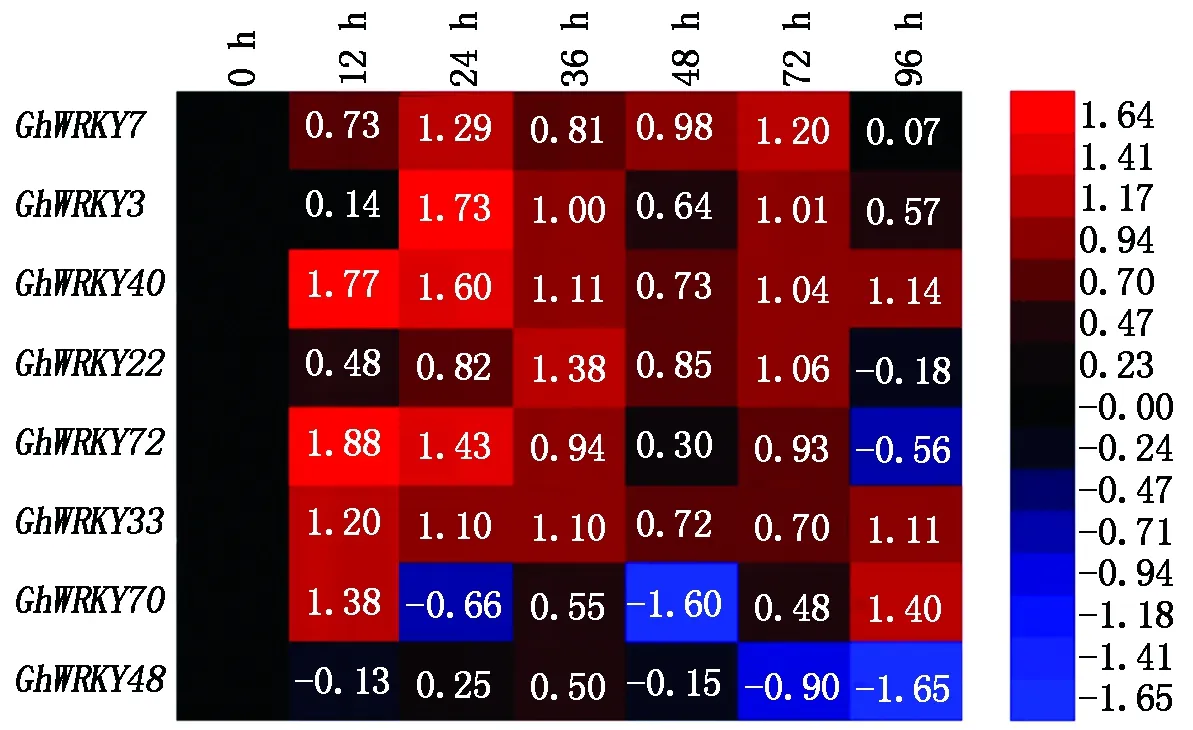
热图的数值为相对表达量进行log2对数转换值,处理0 h相对表达量对数值为0。The log2 value of the relative expression level shown in the heat map,and the value of 0 h treatment level is regarded as 0.
2.3 GhWRKY22组织表达和植物激素诱导表达分析
从上述正调控抗病的5个GhWRKY基因中,挑选GhWRKY22进行深入的抗病功能分析。利用qRT-PCR分析棉花根、茎和叶器官中GhWRKY22基因表达量,结果显示,该基因在棉花根、茎和叶中均有表达,但存在显著差异(图3-A),其在茎中表达量最高,在叶中表达量最低。在图2中已暗示GhWRKY22基因表达可能受到SA和JA抗病信号途径的影响,因此,分别外源使用这2种抗病相关激素处理棉花植株,然后检测GhWRKY22基因的表达情况。图3-B显示,在SA诱导3 h,GhWRKY22基因表达有了显著的升高;而在JA处理后6,12 h,该基因表达呈现显著上升。这些数据表明,GhWRKY22基因表达受到SA和JA信号途径的调控,从而参与棉花的抗病反应。
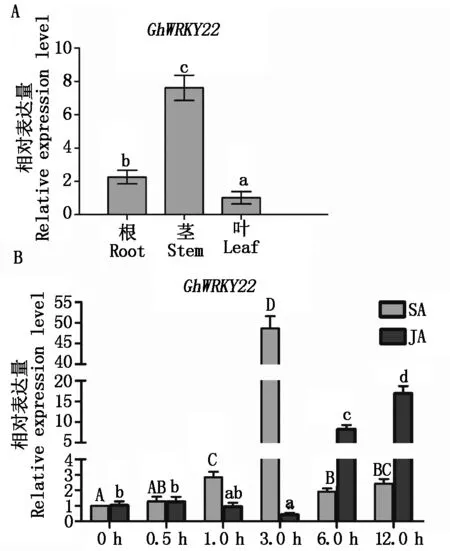
A.GhWRKY22在棉花不同组织中的表达,不同小写字母表示差异达5%(P<0.05);B.SA和JA诱导下GhWRKY22表达情况。不同大写字母表示SA处理下差异性达5%(P<0.05);不同小写字母表示JA处理下差异性达5%(P<0.05)。
2.4 GhWRKY22基因沉默植株的培育
为了进一步分析GhWRKY22抗病功能,利用病毒诱导基因沉默(VIGS)技术来培育GhWRKY22基因沉默植株,分析GhWRKY22基因表达抑制时棉花植株的抗病性。首先,利用NCBI数据库分析比对设计GhWRKY22基因特异靶序列(图4-A、B)。利用BamHⅠ和KpnⅠ双酶切把这个序列插入到烟草脆裂病毒(TRV)载体pYL156中,GhWRKY22构建病毒诱导基因沉默载体pYL-156-GW22(图4-B)。
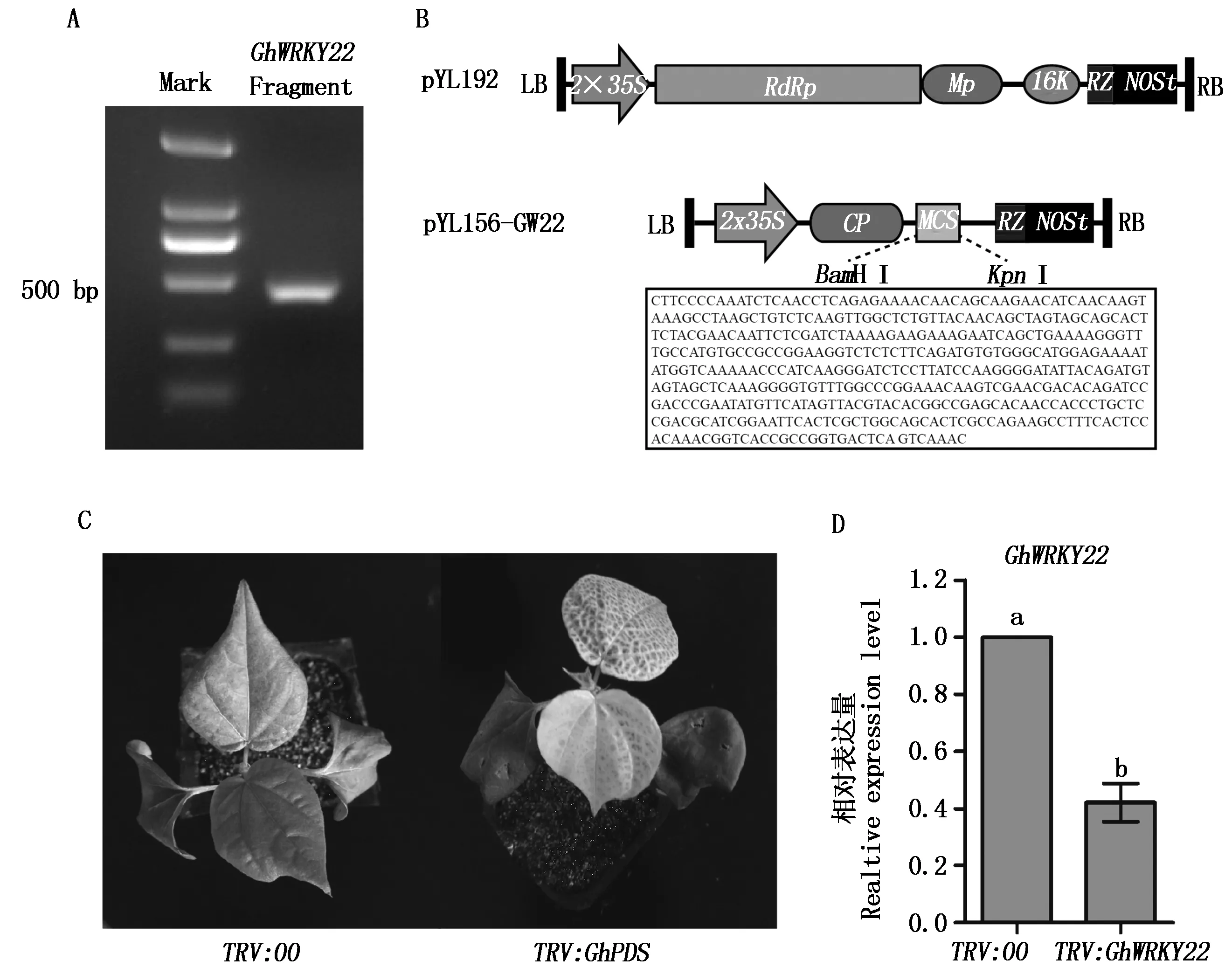
A.GhWRKY22目的片段条带的克隆;B.pYL192辅助载体与pYL-156-GW22沉默载体构建示意图,pYL192载体大小为6.8 kb,其中包含RNA依赖的RNA聚合酶(RNA-dependent RNA polymerase,RdRp)、运动蛋白(Moveprotein,Mp)、16 ku蛋白(16 ku)序列,一个具有自我剪切功能的核酶(Ribozyme,Rz)序列以及强启动子(2×35S) 和NOS终止子序列,pYL-156-GW22载体大小为 10.4 kb,其中含有一个衣壳蛋白(Coatprotein,Cp)序列,一个用于各种目的基因片段构建的多克隆位点(Multiple clone sites,MCS)等;C.GhPDS沉默植株叶片白化表型;D.GhWRKY22基因沉默分析,不同小写字母表示差异达5%(P<0.05)。图5-6同。
当棉花苗的2片子叶完全展开时,进行农杆菌注射浸染。空载体pYL156农杆菌作为浸染负对照,pYL156-PDS农杆菌浸染作为正对照,PDS基因被沉默的植株表现为叶片失绿,一般作为病毒诱导基因沉默的标记基因[22]。接种后的植株继续在培养箱中培养14 d左右,正对照植株呈现叶片失绿白化表型(图4-C),说明TRV系统在棉花中具有系统的感染,并能成功地进行基因表达沉默。此时,GhWRKY22基因沉默植株的幼嫩叶片被取样进行表达量分析。结果表明,GhWRKY22基因在沉默植株中平均表达量显著下降了60%左右(图4-D)。选择GhWRKY22基因表达沉默60%以上的10个植株接种大丽轮枝菌,分析这些沉默植株的抗病反应。
2.5 沉默WRKY22基因植株对大丽轮枝菌的抗性分析
GhWRKY22沉默植株接种黄萎病菌21 d后,发现基因沉默植株表现为对病原菌侵染更敏感。相对于空载体沉默对照植株的发病症状,GhWRKY22沉默植株叶片黄色程度加重,落叶更多;GhWRKY22沉默植株的发病率和发病指数显著高于对照(图5-A-C),发病株率和病情指数分别比对照升高了18百分点,20%,这些数据表明了GhWRKY22基因正调控棉花的抗病反应。为了进一步分析GhWRKY22基因的抗病功能,接菌沉默植株中大丽轮枝菌菌丝生长情况也被检测。首先分析接菌植株主茎断面在培养基上病原菌回复情况,结果显示,GhWRKY22基因沉默植株的病原菌回复率明显高于对照(图5-D)。归纳这些试验结果,可以证明GhWRKY22基因参与棉花对黄萎病的抗性,并且正调控棉花的抗病性。
GhWRKY22基因表达受到SA和JA激素的诱导(图3),因此,GhWRKY22沉默植株对黄萎病的抗性下降,也可能涉及SA和JA信号途径中的基因调控。为了验证这个假设,利用qPCR技术分析SA和JA信号途径中抗病标志基因的表达情况。在GhWRKY22基因沉默植株和对照接种大丽轮枝菌21 d后,分析SA和JA抗病标志基因PR1、PR3、PR4、PR5、PAL1和PDF1.2表达水平。结果表明,基因沉默植株中6个抗病标志基因的表达量显著低于对照植株,其中PR4和PDF1.2的表达量下降最大,仅为对照的3.7%,8.7%。这些结果进一步表明GhWRKY22基因通过SA和JA信号途径参与棉花对大丽轮枝菌的抗性(图6)。

A.GhWRKY22基因沉默植株发病情况;B.发病株率,试验重复3次,共计60个植株;C.病情指数,试验重复3次,共计60个植株;D.接菌植株茎段恢复培养5 d后大丽轮枝菌生长情况。
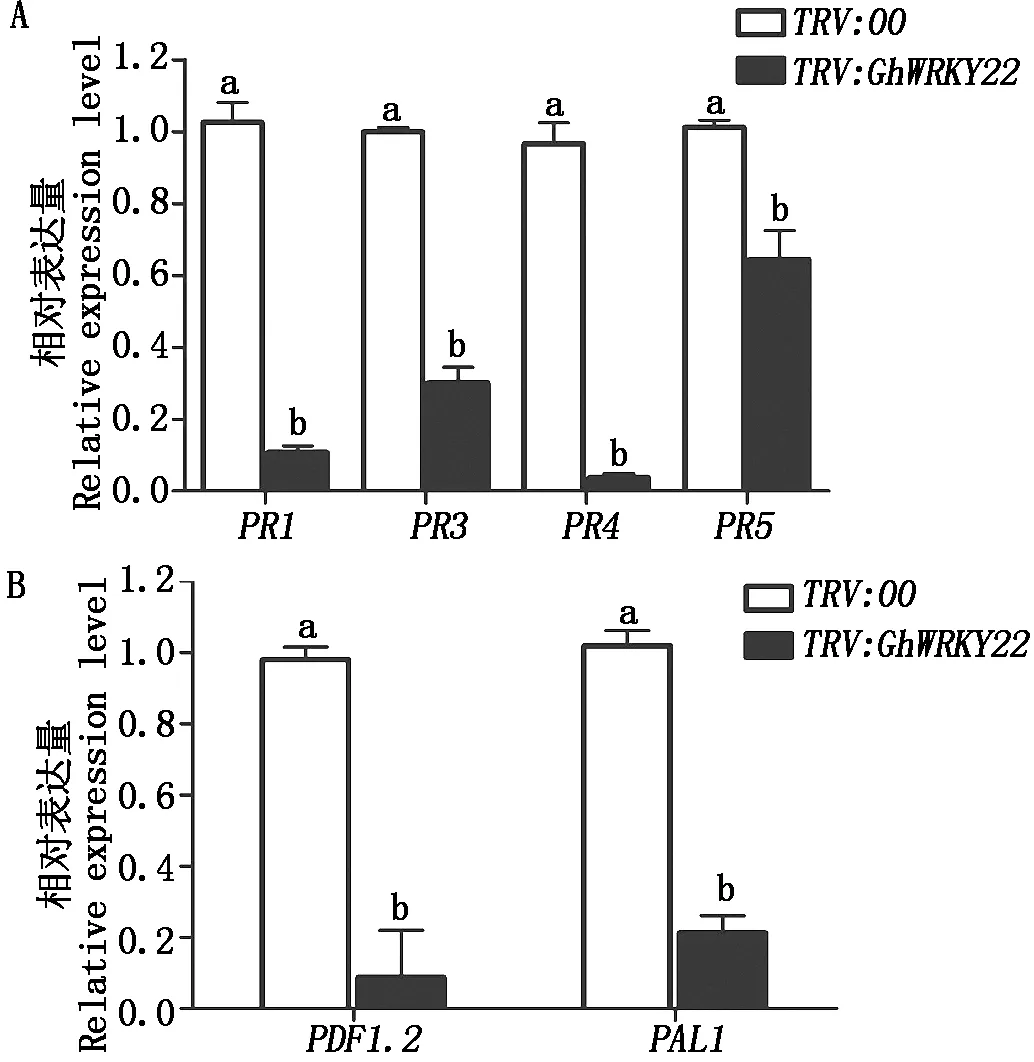
图6 大丽轮枝菌浸染GhWRKY22沉默植株和对照中抗病标志基因的表达分析Fig.6 The relative expression level of defense marker genes in GhWRKY22-silenced plants treated with V. dahliae
3 结论与讨论
植物WRKY基因参与一系列的生理生化过程,包括植物的生长发育和对外界环境的抗性。其中有许多WRKY蛋白参与植物对病原菌抗性,在模式植物拟南芥中已经报道了许多WRKY转录因子参与抗病反应。然而在棉花中只有少数几个WRKY蛋白被报道参与抗病反应,如GhWRKY3[18]、GhWRKY40[19]和 GhWRKY1[20]。相比较于拟南芥,棉花中肯定存在着其他没有鉴定的WRKY蛋白也参与抗病反应。因此,利用比较基因组方法,通过生物信息学技术鉴定了8个与拟南芥直系同源的GhWRKY基因,通过接菌诱导反应显示棉花中这8个WRKY基因均参与大丽轮枝菌的应答反应。并利用基因沉默技术证明了GhWRKY22参与棉花的抗病反应。
本研究结果显示,8个棉花WRKY基因的表达受到大丽轮枝菌的诱导,但是诱导表达的趋势不同,GhWRKY7、GhWRKY72、GhWRKY40等6个WRKY基因表达一直为上调,而GhWRKY48的表达表现为下调,GhWRKY70的表达比较复杂。在棉花中前人已经报道了3个抗病相关WRKY,包括GhWRKY3[18]、GhWRKY40[19]和 GbWRKY1[20]。Guo 等[18]报道了GhWRKY3表达受到多种激素诱导,同时也受到立枯丝核菌(Rhizoctoniasolani)、棉花炭疽菌(Colletotrichumgossypii)和尖孢镰刀菌(Fusariumoxysporum)的诱导。在SA、MeJA和ET处理下,GhWRKY40基因表达上调,正调控茄科植物对雷尔氏菌(Ralstoniasolanacearum)抗性[19]。Li等[20]研究结果显示海岛棉GbWRKY1通过JA信号途径介导棉花抗病和发育的转变,负调控棉花对灰葡萄孢菌(Botrytiscinerea)和大丽轮枝菌抗性。在本研究中的8个直系同源的抗病WRKY基因包含着GhWRKY3和GhWRKY40,都是正调控棉花黄萎病的抗性反应。另外,本研究中有6个WRKY基因正调控棉花大丽轮枝菌应答反应,但同时也存在负调控应答反应GhWRKY48。拟南芥中,利用突变体和过表达植株也证明了WRKY48负调控细菌基础抗性以及PR基因的表达[11];上面论述的海岛棉GhWRKY1也是一个抗病负调控因子;在番茄中,SlWRKY70的转录水平也表现为负调控植株对真菌的抗性[26]。因此,WRKY转录因子参与植物的抗病反应既可以是正向调控也可以是负向调控。
本研究表明,GhWRKY22基因受到大丽轮枝菌浸染的诱导,其表达量随着时间的变化呈现双峰形式的上调,利用VIGS技术确证了GhWRKY22正调控棉花对大丽轮枝菌的抗性。通过激素诱导GhWRKY22防御反应,发现其可能是受到JA和SA交叉信号转导途径的调控,是植物抗病反应中重要调控蛋白,有望成为一个新的棉花抗病育种的候选基因。Zhang等[27]也报道了一个棉花WRKY22基因的抗病功能,这个基因和本研究中的GhWRKY22基因是同源基因,不是拟南芥WRKY22的直系同源基因;然而有趣的是这2个基因都参与棉花对大丽轮枝菌的抗性,它们之间是否存在功能叠加或冗余现象有待深入的研究。在拟南芥中[10],通过野生型与突变体wrky22-1和wrky22-2在淹水处理下比较,发现WRKY22也具有较低的抗病性和先天性免疫特性。此外,Abbruscato等[28]在水稻也报道了OsWRKY22基因位于信号转导回路的交汇点,参与对稻瘟病的抗性。总之,GhWRK22是一个抗病正调控转录因子,可以作为抗病候选基因用于棉花的抗病品种培育。
