水杨酸诱导黄瓜PCD的鉴定及相关基因的表达分析
2019-01-09王丹丹池春玉金晓霞丁国华
陈 冲,刘 双,王丹丹,池春玉,2,朱 宏,2,金晓霞,2,丁国华,2
(1.哈尔滨师范大学 生命科学与技术学院,黑龙江 哈尔滨 150025;2.植物生物学黑龙江省高校重点实验室,黑龙江 哈尔滨 150025)
水杨酸(Salicylic acid,SA)是植物内源性信号分子,参与调节植物许多生理过程[1-2]。外施SA可以提高植物对盐[3]、高热[4]、低温[5]、干旱[6]、紫外线[7]、重金属[8]等非生物胁迫的耐受性。
SA在提高植物抗病性方面的作用尤其受到人们的关注[9-10]。SA是植物受到病原菌侵害时发生超敏反应(Hypersensitive response,HR)以及形成系统获得性抗性(Systemic acquired resistance,SAR)所需要的内源信号分子[11-12],阻断SA的生成或促进SA的分解,都会出现HR的延缓和病斑扩散的现象[13]。有人对辣椒等植物外源施用SA,也有效提高了植株对疾病的抗性和病程相关基因(PR)的表达[14]。
有很多研究证实,SA介导的HR是一个程序性细胞死亡(Programmed cell death,PCD)过程[15-16]。植物发生HR时,常伴随着特异性信号分子ROS和SA的积累以及PR的表达,激发邻近组织的特异性防御反应和植株的SAR[12]。线粒体在PCD中扮演重要角色[17-18]:有研究表明,线粒体通透性转运孔(Mitochondria permeability transition pore,MPTP)的开放是PCD中发生的重要信号之一[19]。MPTP的开放引起线粒体膜电位下降,进一步导致氧化磷酸化作用的解偶联,膜内外离子浓度趋于平衡,进而导致线粒体外膜被破坏和细胞色素C(Cytochrome C,Cyt C)的释放,释放到胞质内的Cyt C可以激活Caspase等下游关键酶类,使信号级联放大而触发PCD[20-21]。但近年来人们开始注意叶绿体在PCD中的作用[22]。早在2002年就有报道,氰化物(CN)在光刺激下引发含叶绿体的豌豆果皮细胞的PCD过程,而不含叶绿体的表皮细胞却不受影响[22-24]。活性氧(Reactive oxygen species,ROS)具有信号和触发植物PCD的作用,而叶绿体也是ROS的重要来源,因此,叶绿体对植物PCD具有重要作用[25]。
被首先关注的就是ROS的作用。有人发现,叶绿体通过光合电子传递和叶绿素生物合成途径产生ROS,导致核心天线复合体降解,并激活 Chloroplastic Executer 1、2 蛋白,继而将信号传递到细胞核,激活细胞死亡相关基因的转录[26-27]。
尽管植物基因组上没有找到编码caspase的基因,但还是找到了一些具有caspase活性的蛋白酶[31-32],一些定位在叶绿体的蛋白酶和基因开始被关注,其中有的有caspase活性,与PCD关系密切[33-36]。定位在叶绿体膜上的Cid和Lrg蛋白被发现可以执行类caspase活性,在细胞PCD中起作用[37]。
为了获取更多的与PCD相关的基因信息,植物生物学黑龙江省高校重点实验室采用数字转录表达谱技术获得了1 759个SA诱导表达的基因[38],通过基因注释筛选出15个与叶绿体相关的基因。为了揭示在SA诱导PCD中叶绿体所起的介导作用,本研究在对SA处理前后的黄瓜进行基于活性氧(ROS)原位检测、台盼蓝(Trypan blue)染色、碘化丙啶(PI)和荧光素二乙酸酯(FDA)染色、转移酶介导的dUTP缺口末端标记测定法(TUNEL)等PCD鉴定,对15个筛选得到的基因进行RT-PCR、实时荧光定量PCR的表达分析,为研究这些基因的功能和介导PCD中的作用奠定基础。
1 材料和方法
1.1 材料培养
以东农649为试验材料,在光照培养箱内培养,设置为10 h黑暗、14 h 66%光照的培养条件,温度分别为24,28 ℃。
1.2 ROS原位检测

1.3 Trypan blue染色法
取材方法同上,染色方法采用Stone和Bowling的试验方法[40-41]。把材料浸到Trypan blue染色液中,100 ℃水浴2~3 min后,室温(25 ℃)静置1 h,用2.5 g/mL的水合氯醛溶液将材料煮沸20 min,在此过程中需数次更换水合氯醛,至材料完全褪色为止,取材料至50%甘油(V/V)中贮存和装片,在体视显微镜下观察。若染色结果为阳性,则材料经处理的部分呈蓝色。
1.4 PI和FDA染色
取经处理材料叶片下表皮细胞,清洗材料至SA完全去除,用FDA染液染色材料30 min,用双蒸水清洗材料,再用PI染色10 min,冲洗后用50%甘油(V/V)装片进行观察,荧光素激发波长为488 nm,发射波长为530 nm。PI与FDA的协同使用,分别对死细胞和活细胞染色。
1.5 TUNEL法检测PCD
采用Roche公司的In Situ Cell Death Detection Kit(11684817910)试剂盒,对SA处理后的黄瓜表皮细胞和黄瓜叶片进行TUNEL显色法检测,具体步骤见说明书。
当我们抛开“权能”客观方面的含义,仅把“权能”看成是一种积极的能动的精神时,我们发现增能理论的核心观念“权能”与陆九渊心学的核心观念“本心”具有极大的相似性。
1.6 黄瓜超敏反应相关基因的RT-PCR和qPCR表达分析
黄瓜幼苗处于四叶期时,在叶片每隔3 cm滴加50 μL 10 mmol/L SA,分别在0,3,12,24 h这4个时间段取材并用液氮冷冻,在-80 ℃冰箱保存。分别进行半定量RT-PCR和实时荧光定量PCR测定,检测SA处理下,15个超敏反应相关基因在黄瓜叶片中的表达水平。
2 结果与分析
2.1 黄瓜PCD的鉴定
2.1.1 SA处理黄瓜叶片的形态变化 如图1-A所示,用10 mmol/L SA处理黄瓜幼苗叶片后,与对照组叶片(0 h)相比,处理组叶片会出现变薄的趋势,并随着处理时间的加长,叶片的变薄程度也随之增加。3 h时处理组叶片出现变薄趋势,且在6,9,12 h变薄程度明显,并在24 h达到极值。试验结果表明,处理3 h时叶片细胞开始发生PCD,并在24 h达到最大值。
2.1.2 ROS的定位 如图1-B、C所示:用SA处理黄瓜幼苗叶片后,3 h叶片出现了棕色和蓝色,说明在此时已经有ROS的积累。随着处理时间的增加,处理部位的颜色逐渐变深,ROS增多。24 h与6,9 h相比,颜色进一步加深,在处理部位周围也出现了棕色和蓝色的积累。结果表明,在SA诱导黄瓜幼苗叶片细胞死亡的过程中,伴随有ROS的暴发。
2.1.3 Trypan blue染色 如图1-D所示,与对照组叶片相比,SA处理黄瓜幼苗叶片3 h后,染色区域出现浅蓝色;6 h与3 h相比,染色区域的大小和颜色的深浅变化不大,但处理9 h后,染色区域的颜色逐渐变深;处理12,24 h后,染色区域的颜色加深呈现深蓝色,且染色区域开始向未经SA处理的区域扩散。试验结果表明,随着SA处理黄瓜叶片的时间增加,死亡的细胞也逐渐增多。

0 h.对照;3,6,9,12,24 h.10 mmol/L SA处理时长。A.叶片形态;B.DAB染色; C.NBT染色;D.Trypan blue染色。
2.1.4 PI和FDA染色结果 如图2所示,对照组表皮细胞在荧光显微镜下,无红色荧光出现,说明对照组表皮细胞并未被PI染色,即对照组表皮细胞并没有死亡细胞的出现;FDA染色可见巨量的亮绿色荧光(图2-A),此时的细胞都是活细胞。而经过 SA处理1,3,5 h后,荧光显微镜下的红色荧光逐渐增多,绿色荧光逐渐减少(图2-B-D)。结果表明,随着SA处理时间的延长,黄瓜叶片表皮细胞的死亡比例呈现一个逐渐上升的趋势。

A. 未处理的黄瓜表皮细胞;B. SA处理1 h的黄瓜表皮细胞;C. SA处理3 h的黄瓜表皮细胞;D. SA处理5 h的黄瓜表皮细胞。
2.1.5 TUNEL法鉴定PCD 如图3所示,对照组细胞A0没有颜色出现,用苏木精复染(B0)后,细胞核多呈现卵形或球形,说明此时没有细胞死亡;使用SA处理3 h(A3)后,有少量的细胞核的颜色发生变化,细胞核变形,这是细胞程序性死亡前期细胞核的典型表现;随着SA处理时间的增加(A6、A9、A12),细胞核颜色变化明显,出现密集的阳性斑点,经过苏木精复染(B6、B9、B12)的细胞核形状发生明显的改变和边缘化,这是细胞程序性死亡加剧的表现。

A. DAB染色;B. 苏木精复染;0 h.对照;3,6,9,12 h分别是SA处理黄瓜叶片的时间。

表1 SA诱导黄瓜叶片差异基因中叶绿体相关基因Tab.1 SA induces chloroplast related genes in cucumber leaf differential genes
2.2 SA诱导黄瓜PCD相关基因的筛选
分析Illumina高通量测序技术获得的差异表达基因片段[38],从1 759个差异表达基因DEGs中,用PCR验证的方法筛选出15个注释中带有chloroplast的基因进行表达分析(表1)。
2.3 SA诱导黄瓜叶绿体PCD相关DEGs的表达分析
图4是用qRT-PCR的方法检测15个基因在SA处理下不同时间段的表达量变化。有12个基因都表现先上调后下调(A4、A5、A6、B3、C3、D1、E3、E4、E6、E7、E8和G3),其中A4、A5、B3、C3、E4和E7在SA处理3 h时表达量最高,之后下降;A6、D1、E3、E6、E8和G3在SA处理12 h时表达量最高之后下降。D6和A2属于表达逐步下调的基因。F7的表达在处理第12 h降至最低,至24 h表达又恢复至对照水平。

图4 SA处理15个基因的qRT-PCR表达分析Fig.4 Real time quantitative RT-PCR expression analysis of fifteen genes under SA treatment
按相对表达量进行分析,与处理前相比,表达量上调不足2倍的只有A5、C3和E4等基因,其他基因都对SA有强烈的表达响应,其中A6、D1、E6的最高表达量比处理前分别高出9,6,10倍。所有基因的上调表达在最高表达量上都不持续,之后的测定都显示表达量已下调直至接近处理前水平,甚至低于处理前(A6、G3)。
基因F7的表达是先下降后上升,其最低表达量与处理前相比差2.5倍,即受SA的诱导作用不很明显。2个下调表达的基因A2和D6受SA影响比较明显,与处理前相比下调程度都达到5倍。
3 讨论
植物细胞程序性死亡(PCD)在植物系统生长、器官形成和对抗逆境中都有重要意义[42]。而ROS和SA作为植物PCD的重要内源信号分子,植物在响应胁迫时常伴随着ROS的积累和爆发[43]。
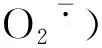
对选定的15个黄瓜基因进行荧光定量PCR表达分析,响应SA诱导作用表现出上调表达的基因有:类囊体加工肽酶1基因(B3)、6-磷酸葡萄糖酸脱氢酶基因(D1)、果糖1,6-二磷酸醛缩酶3基因(E3)、α-葡聚糖-水二激酶1基因(E6)和丙二烯氧化物环化酶4基因(E8);下调表达的基因有:甜菜碱脱氢酶1基因(A2)、钙敏感受体基因(A4)、脱镁叶绿酸a单加氧酶基因(A5)、叶绿素还原酶1基因(A6)、σ因子结合蛋白1基因(C3)、NADPH-原叶绿酸酯还原酶基因(D6)、2-琥珀苯甲酸-辅酶A连接酶基因(F7)、ATP依赖的锌金属蛋白酶基因(E7)、烟酰胺腺嘌呤二核苷酸转运子基因(E4)、胱硫醚β合成酶(CBS)蛋白基因(G3)。
查阅文献并根据相对表达量的变化,推测了15个基因的功能并分为以下6类:叶绿素合成降解相关基因(PaO、NYC1、NPR);植物抗逆性相关基因(6-PGDH、FtsH1、NDT1);调控SA和JA诱导的植物防卫反应相关基因(AOC4、SIB1);提高植物胁迫耐受性相关基因(BADH1);植物光合作用相关基因(MenE、FBA3);调节植物发育相关基因(CBSX1、GWD1、TPP1、CAS)[46]。
SIB1属核编码的植物特有的抗病原体相关基因,Narusaka等[47]发现拟南芥基因SA的诱导,且在SIBI超表达株系中发现与ROS积累相关的基因转录水平提高,本试验中SIB1表达呈先上升后下降的趋势,印证了该基因参与SA诱导提高植物免疫力的信号途径[47],推测其可能连接叶绿体功能并参与植物防御系统[48]。
FtsH1在SA处理3 h先表达上调,12 h后表达量恢复正常并有所下降,这个结果与烟草受病毒侵染后FtsH1表达下调加速超敏反应是一致的[49]。FtsH1在拟南芥中参与类囊体蛋白的降解,控制蛋白质的质量平衡[50],降解光系统Ⅱ(PS Ⅱ)修复中光损伤反应中心D1蛋白[51]和一些组装PS Ⅱ剩余的蛋白亚基[52]。
PaO在本试验中表达量呈应激性升高,并在之后的时间呈下降的趋势,认为PaO在处理早期时发生应激反应,加速了光合作用,随后下调使光合作用后续产物脱镁叶绿酸a积累,终止光合作用,执行使细胞加速死亡的功能;NYC1的表达量在本试验中呈先上升后下降的趋势,认为NYC1通过诱导LHCⅡ的降解来增加叶绿素a的合成,参与植物光合作用进程,进而参与调控SA诱导PCD的进程。PaO基因受SA诱导下调表达,促进HR,其原因是缺失该酶导致光毒性的脱镁叶绿酸a积累引起光依赖的PCD[53-54]。Horie等[55]研究表明,拟南芥缺失该基因后,会在植物发育期间表现出滞绿表型。光系统Ⅱ的捕光叶绿素a/b蛋白复合物(LHCⅡ)是色素蛋白复合物,结合叶绿素a和叶绿素b并参与调节PS Ⅰ 和PSⅡ能量的分配。叶绿素b是LHCⅡ的主要色素,在NYC1突变体株系衰老期间,叶绿素b的降解被抑制导致LHCⅡ选择性的保留,这表明叶绿素b还原酶参与LHCⅡ降解起始进程。
AOC4是茉莉酸(JA)合成的关键酶,SA处理后,苹果AOC1基因的表达量上升,JA含量也随之增加[56]。喜树AOC基因在细菌中表达后,提高了细菌对盐的耐受性[51]。本试验中AOC4上调表达的结果与提高抗性相一致。NDT1在烟酰胺腺嘌呤二核苷酸(NDA)的运输过程中起重要作用,植物受到病原体侵染时,NAD调控某些次生代谢产物来产生抗性诱导植物发生超敏反应[43]。本试验中NDT1上调,认为NDT1参与了SA提高植物抗性的信号途径。
