杂草对合成生长素类除草剂的抗性
2019-01-05编译
叶 萱 编译
(上海市农药研究所,上海 200032)
合成生长素类除草剂(SAH,HRAC分类体系的O 组)模拟了天然植物激素吲哚-3-乙酸(IAA)的作用。第1个这种作用机制的除草剂2,4-滴已被广泛使用70多年。SAH主要用于选择性防治禾本科作物田中的阔叶杂草,但二氯喹啉酸和氯氟吡啶酯防治一些禾本科和沙草科杂草。SAH被分为7种:⑴苯氧羧酸类;⑵ 苯甲酸类;⑶ 吡啶羧酸类;⑷ 吡啶氧羧酸类;⑸ 喹啉羧酸类;⑹ 嘧啶羧酸类和⑺芳基吡啶甲酸类(arylpicolinate)(表1)。每一种都具有独特的化学结构(图1)。自1945年2,4-滴引入后SAH就一直被商业化使用到现在,在2018年氯氟吡啶酯引入(表 1)。2,4-滴引入用于农业给杂草管理带来了革命,持续性创新发现和开发了数个新颖的SAH。
SAH的全球施用面积仅次于 ALS抑制剂(508×106hm2)和 EPSP 合酶抑制剂(477×106hm2)除草剂,位列第三(366×106hm2)(信息来自陶氏益农,2014年)。除草剂2,4-滴的全球使用面积为161.7×106hm2,是使用最广泛的SAH,其次依次为麦草畏(50.0×106hm2)和 2-甲基-4-氯苯氧乙酸(MCPA)(31.3×106hm2)(图 3)。

表1 不同种类合成生长素类除草剂
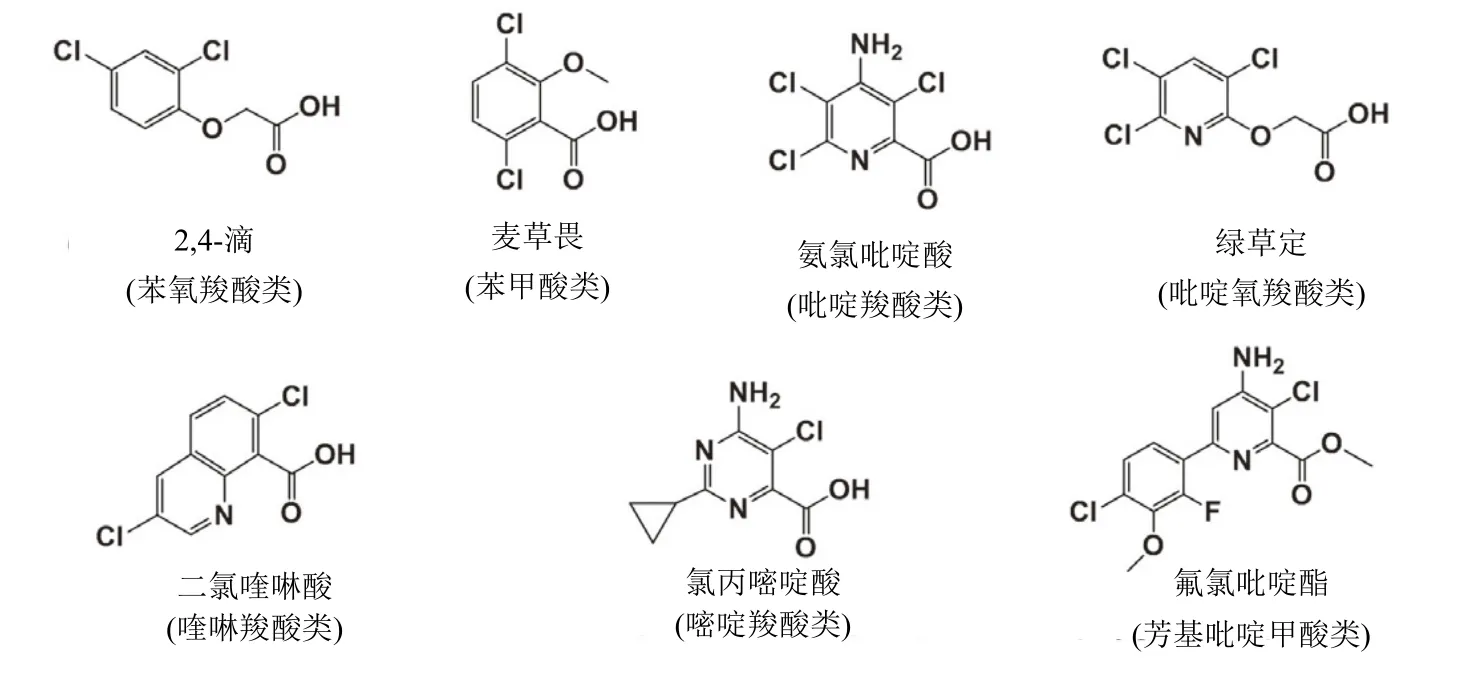
图1 7种类别合成生长素除草剂代表性产品结构
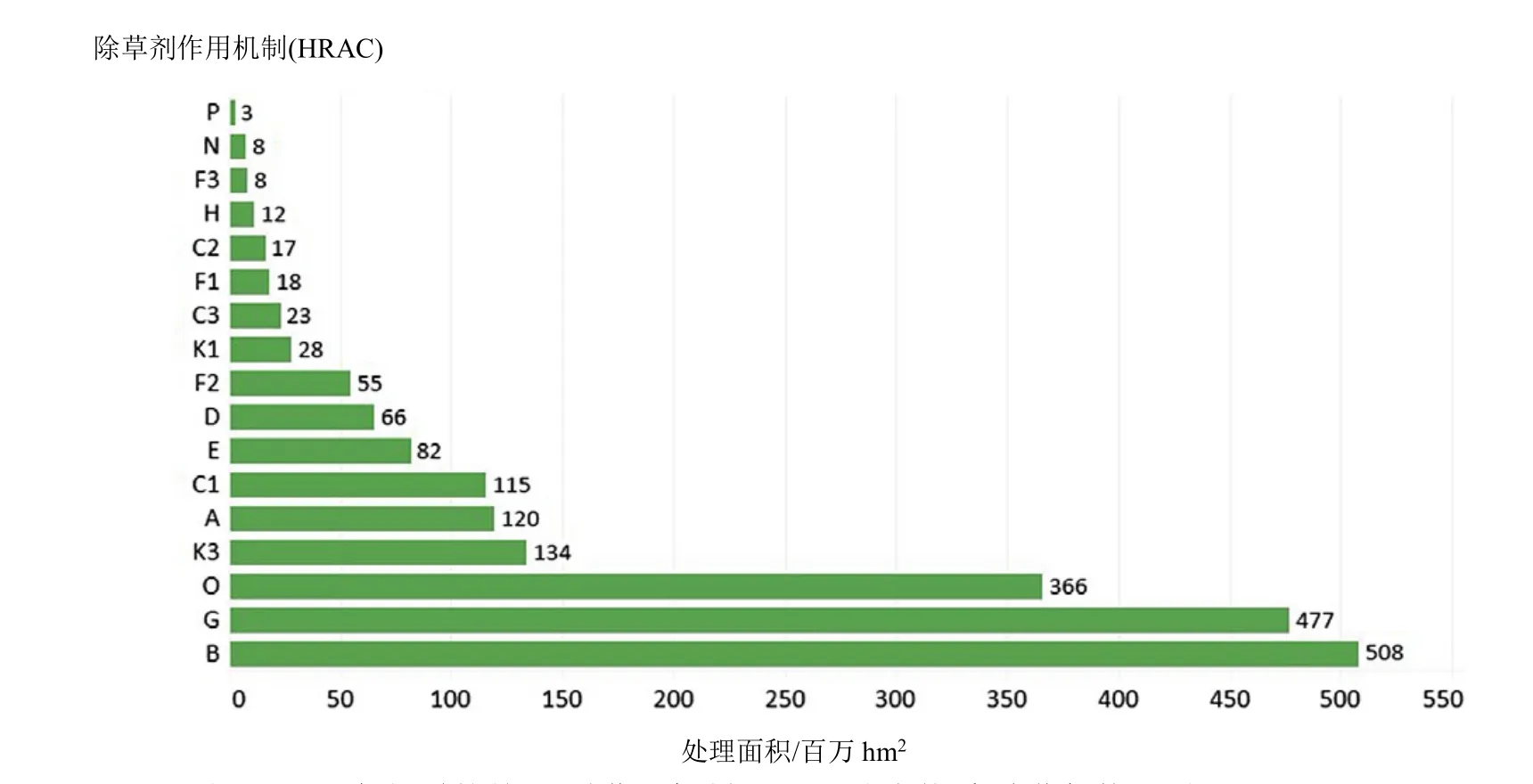
图2 2014年报道的前17种作用机制(HRAC分类体系)除草剂使用面积(×106 hm2)
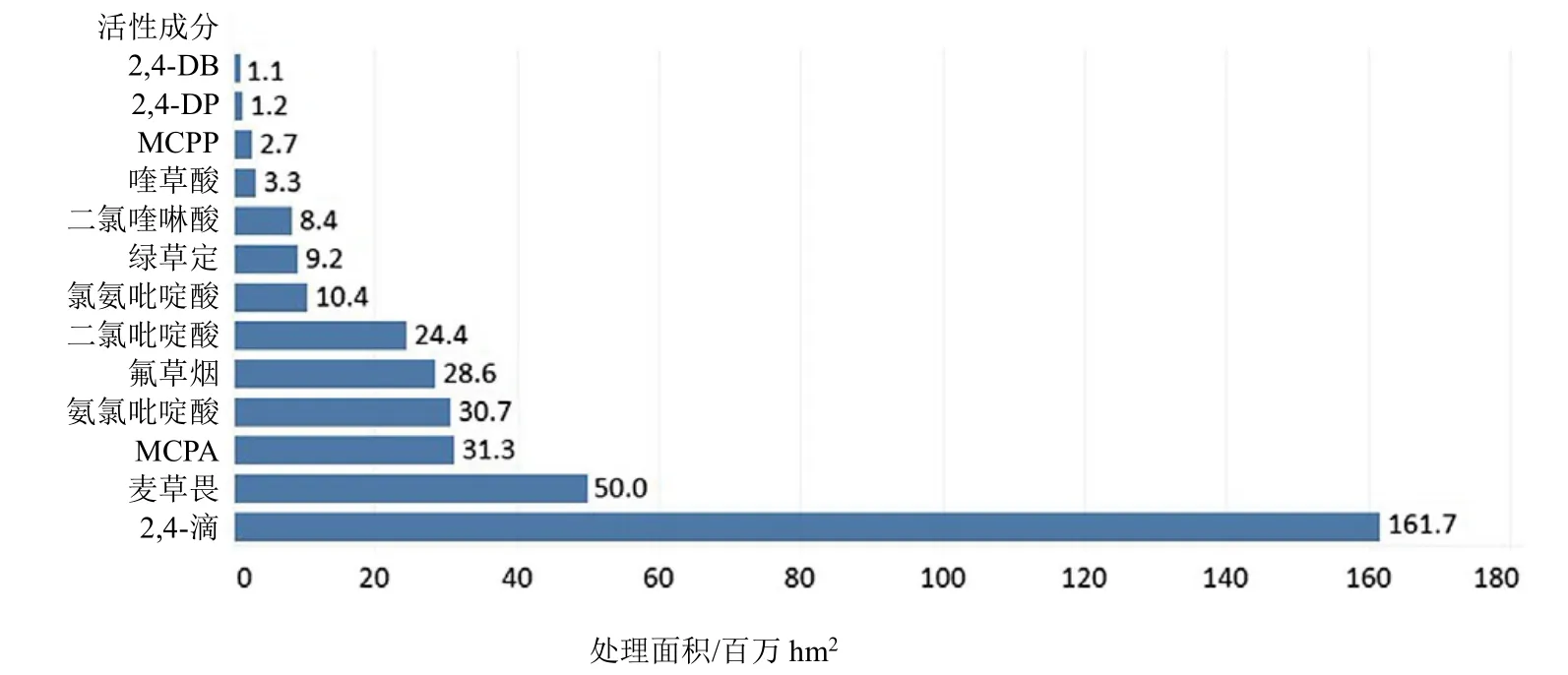
图3 2014年报道的特定合成生长素类除草剂活性成分施用面积(×106 hm2)
为了持续应用 SAH,掌握杂草对SAH的抗性机制、抗性选择和进化知识非常重要。在2017年5月在美国科罗拉多州的丹佛举办的“全球除草剂抗性挑战”会议期间,进行了“杂草对SAH抗性”的专题讨论会。讨论会的主题有杂草品种对SAH抗性的进化;熟知的杂草品种对 SAH抗性进化的机制;SAH的感应、运输和代谢;SAH发现的最近创新;耐SAH作物的抗性管理。
1 杂草品种对SAH的抗性
在1957年,首例2,4-滴抗性被报道,即在夏威夷节节草(Commelina diffusa)和在加拿大胡罗卜(Daucus carota L.)对2,4-滴产生了抗性。根据“国际抗性杂草调查”,现在有36种抗SAH的杂草品种(30种阔叶杂草、5种禾本科杂草和1种类似禾本科杂草)(图4)。包括止血马塘(Digitaria ischaemum)和4种稗属杂草[稗草(Echinochloa crus-galli)、孔雀稗(Echinochloa crus-pavonis)、西来稗(Echinochloa zelayensis)和芒稗(Echinochloa colona)]的5种禾本科杂草已对二氯喹啉酸产生了抗性。有人提出其为氰化物介导的抗性机制,不同于SAH的。
对SAH产生抗性的经济重要性杂草有澳大利亚抗2,4-滴和抗MCPA的野萝卜(Raphanus raphanistrum)、欧洲抗苯氧除草剂的虞美人(Papaver rhoeas),加拿大和美国抗麦草畏的地肤(Kochia scoparia),美国抗 2,4-滴、麦草畏和MCPA的刺莴苣(Lactuca serriola)。此外,内布拉斯加州和伊利诺斯州的长芒苋(Amaranthus tuberculatus)生物型和阿根廷的绿穗苋(Amaranthus hybridus )被确定对一些SAH有抗性。经济重要性稍差的抗性杂草有加拿大和美国抗2,4-滴的胡罗卜,新西兰抗 2,4-滴的麝香飞廉(Carduus nutans)和意大利蓟(Carduus pycnocephalus),以及在加拿大对SAH多抗的野芥(Sinapis arvensis L.)和抗二氯喹啉酸的猪殃殃(Galium spurium)。
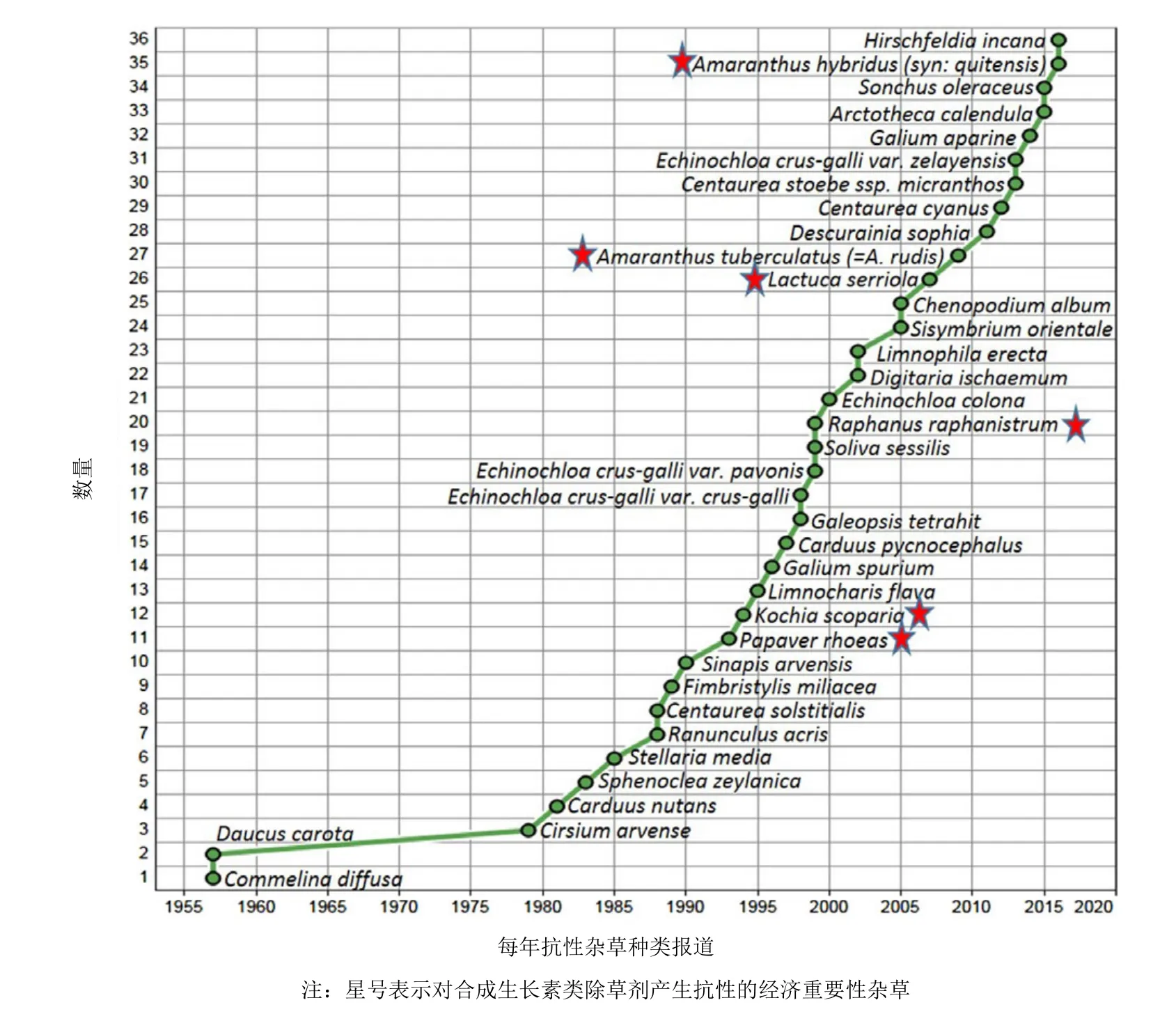
图4 到2017年已报道对合成生长素类除草剂产生抗性的杂草品种
与其他作用机制除草剂特别是乙酰基辅酶A羧化酶(ACCase)和乙酰乳酸合酶(ALS)抑制型除草剂相比,考虑到广泛使用SAH所形成的选择压程度,其抗性发生程度较低(图5)。虽然SAH使用的时间要长于其他作用机制的除草剂,但相对来说没有对农业生产造成广泛的不利影响的抗 SAH杂草案例(图 4)。
SAH抗性发生低的原因可能有以下几种:⑴ 这些除草剂具有潜在的多个作用位点;⑵ 一些抗性事例是由隐性基因所致,其扩展比显性性状更慢;⑶在除草剂和作物竞争存在的情况下,抗性基因型的适合度降低。杂草交互抗性的发生可能是管理策略开发的1个挑战。更完全地了解SAH的抗性机制应该能改进抗性管理的措施和更好地应用这些除草剂。
2 一些杂草品种对SAH的抗性机制
2.1 虞美人
虞美人是欧洲冬季谷物中最常见的阔叶杂草。此杂草品种专性异花授粉,种子产量高,种子库具有高度持久性,适合种子萌发的时间长,故难以防治。随着除草剂抗性的出现和扩展,虞美人正成为更难防治的杂草,特别是在欧洲的南部。最近10年在西班牙、法国和希腊,报道出现抗苯氧羧酸类(2,4-滴和 MCPA)和抗乙酰乳酸合酶抑制剂(苯磺隆)的虞美人生物型。在西班牙,也可能存在对其他苯氧羧酸类、苯甲酸类或吡啶羧酸类有交互抗性的生物型。很少有人研究虞美人对SAH的抗性机制和基因。
抗性植物中 2,4-滴迁移的缺少可能是抗性产生的原因。此外,用2,4-滴处理的敏感植物产生的乙烯是抗性植物的4~8倍。2,4-滴在抗性植物中可能没有到达核蛋白受体复合体,压制了生长素反应性基因,其中一些基因负责乙烯的产生。乙烯的积累可能抑制光合作用,产生H2O2和反应性氧,导致植物死亡。
将2,4-滴用于迁移性下降的抗性虞美人种群48 h后,在植物的嫩芽和根中检测到 2,4-滴的代谢物。在处理后的 168 h,没有检测到 2,4-滴,只检测到HPLC保留时间显示为羟基化的2,3-滴和2,5-滴代谢物。用细胞色素P450抑制剂马拉硫磷处理,能使基因型从抗性转为敏感型。这表明细胞色素P450增加了这些抗性种群对 2,4-滴的降解。迁移性降低和除草剂降解的增加可能都是虞美人产生抗性的原因,但哪个是主要机制仍未知。一般在韧皮部的移动性比 2,4-滴更小的极性代谢物能够降低迁移性。其他人发现在 2,4-滴降解前其迁移性就降低了。分化代谢、乙烯反应和迁移性可能是相互依赖或独立机制,确切情况还需进一步研究。
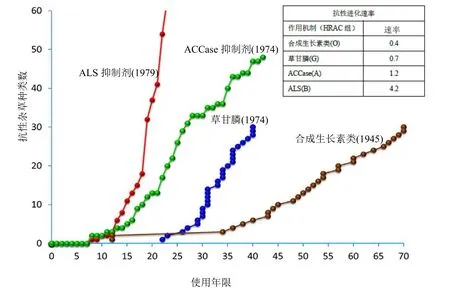
图5 自引入后到2017年期间ALS抑制剂、ACCase抑制剂、草甘膦和合成生长素类除草剂的抗性杂草品种的数量和杂草抗性相对发展率排序
有能够降解甲氧咪草烟的对ALS抑制型和苯氧羧酸类除草剂有抗性的虞美人生物型。是否相同的细胞色素P450降解这些作用机制的除草剂,或是否其他酶参与其中,这些仍未知。发现除草剂 2,4-滴能诱导敏感和抗性虞美人表达糖苷水解酶(GH3)和谷胱甘肽S-转移酶(GST3),因此这些酶表达的增加好像与 2,4-滴的抗性没有关系。需要确定虞美人抗除草剂的基因和更充分地了解其抗性机制。
2.2 地肤
地肤为侵染北美大平原作物地和非作物地的一年生夏季杂草。它出现于早春。雌蕊先熟生物学确保了高水平的远交,导致高的遗传变异性。成熟的种子没有硬的外壳,很少休眠。地肤对莠去津、麦草畏、草甘膦和数种ALS抑制型除草剂产生了抗性。
抗麦草畏地肤被首次发现于20世纪90年代的科罗拉多州和内布拉斯加州。到2016年,抗除草剂生物型已普遍,特别是在科罗拉多州和堪萨斯州的休耕小麦田中。到目前为止,在科罗拉多州还没有发现对氟草烟产生交互抗性,虽然报道蒙大拿州有。在25年中使用先进的选择技术,培育了抗性地肤品系 9425,与敏感品系 7710相比其对麦草畏的抗性增加了30倍。性状为显性或半显性。与敏感品系相比 9425品种对麦草畏的迁移性降低了。RNA-序列表明品系 9425体内影响生长素运输的一组独特基因被下调了,这可能是导致抗性植物中麦草畏迁移性下降的原因。
已发现少量的抗氟草烟地肤种群。虽然氟草烟是有效的可靠的防治各种环境中地肤的除草剂,但由于其杂草防治谱窄,不被看作是“可独立使用”的产品。由于地肤对其他除草剂的抗性广泛,而用氟草烟防治地肤日益增加,管理地肤对氟草烟抗性的进化将非常重要。
2.3 刺莴苣
刺莴苣是菊科一年生、冬性一年生或二年生杂草,是美国西北部的一大问题杂草。此植物具有深的主根,能在干旱条件下存活。刺莴苣除了与作物竞争水分、空间和营养外,其产生的黏白乳胶能堵塞收获设备,增加谷物湿度。在收获过程中与谷物混合的花蕾难于去除,降低了谷物的品质。
美国西北部种植的约80%小麦在生长过程中至少被施用1种SAH 1次。在2004年夏天在美国华盛顿普尔曼附近,应用840 g/hm2草甘膦和540 g/hm22,4-滴 2次后刺莴苣还能够存活。此外调查发现对2,4-滴的差异反应可遗传,防治失败不是草甘膦抗性所致。在被处理后3周抗性植物从花冠再次长出的部分被用 2,4-滴处理后 2~3周受到伤害。抗性水平是敏感生物型的9~10倍。此2,4-滴抗性生物型似乎对苯氧羧酸类和苯甲酸类包括MCPA和麦草畏在内的其他SAH已发展交互抗性。
刺莴苣对 2,4-滴抗性的遗传性受单独共显性基因控制。进行了环标记的14C-2,4-滴生物动力学研究来了解2,4-滴抗性的机制。在应用后96 h,抗性生物型吸收了较少的 2,4-滴,与敏感生物型相比,处理叶中有较多的放射活性。发现在处理的抗性和敏感生物型植物叶或花冠中2,4-滴的代谢率没有差异。
在处理后前4 d,敏感生物型的植物生长受到了刺激,抗性生物型没有。4 d后,敏感生物型的生长几乎停止,而抗性生物型继续生长。抗性生物型持续生长可能是生长素信号传导反应发生改变所致的降解下降引起。也许,抗性生物型体内生长素信号传导的改变和正常生长的维持可能已降低了抗性刺莴苣生物型对 2,4-滴的吸收和迁移。还没有测定基因精确的作用。转录物组学和靶标基因表达研究可能有助于确定引起抗 2,4-滴刺莴苣生长素信号传导反应改变的潜在基因。
2.4 野萝卜
野萝卜是澳大利亚南部种植系统中最成问题的双子叶杂草,每年给种植者造成的作物产量损失至少达5700万澳元,增加了杂草防除费用。在澳大利亚西部,此杂草已对ALS抑制型除草剂的磺酰脲类和咪唑啉酮类产生了广泛的抗性,种植者要依赖SAH特别是2,4-滴来防除。在1999年,首次确定野萝卜在田间对 2,4-滴发展了抗性的事例。随后在2003、2010和2015年进行的随机杂草种群调查表明对2,4-滴产生抗性的植物种群比例从2003年的60%增加到2010年的74%。在2015年抗性水平保持没变。
在2001和2002年从田间收集的2个野萝卜种群的表型特征表明与敏感种群相比抗性增加了约20倍。赋予这2个种群2,4-滴抗性的性状为细胞核遗传和不完全显性。14C-标记的2,4-滴从2002年种群和 2010年收集的种群的被处理叶中迁移出的能力很弱。对敏感植物应用生长素外排抑制剂1-N-萘基酞氨酸能模仿 2,4-滴迁移受限,在抗性植物中 2,4-滴迁移受限可能是由于ATP-结合盒型B型(ABCB)长距离生长素外排运输蛋白功能损失所致。野萝卜的主要抗性机制好像是2,4-滴迁移的下降。
从1999、2001和2002年种群收集的2,4-滴抗性生物型也对MCPA有抗性。对1999年种群中抗MCPA生物型的研究表明其抗性遗传方式相似于抗2,4-滴生物型的。在测定苗根延长的试验中,2002年种群对2,4-滴、MCPA、2甲4氯丙酸、麦草畏和1-萘乙酸有抗性,而2010年种群只对苯氧乙酸、2,4-滴和MCPA有抗性。这表明抗性生物型间存在生长素感知和/或信号传导的差异。对 2002年种群进行的全基因组转录组学研究表明生长素诱导的转录抑制因子和防御基因迅速下调,此现象在敏感种群中没有发现。这可能是第二种抗性机制,因为一些对2,4-滴具有高度抗性的种群如敏感植物一样能有效地从被处理叶中迁移出14C-标记的2,4-滴。
用推荐剂量 2,4-滴处理的和小麦竞争生长的植物与用 2,4-滴处理但没有竞争生长的植物相比,其生物量减少了75%。这可能有助于确定交互抗性水平和除草剂迁移性变化,和/或抗性基因表达水平不同的植物种群在没有进行除草剂处理的情况下的适合度。用此信息能够识别适宜的非化学措施,如早期作物种植,应用高竞争性的作物品种,这有助于更有效的抗性管理策略。
有一些化合物能够阻碍基因或基因产品改变抗性植物对 SAH的反应或再次激活除草剂的正常迁移能力,发现这些替代性分子可能延长SAH防治作物田中野萝卜的时间。到目前为止,没有发现抗性野萝卜对 2,4-滴解毒代谢的证据(细胞色素 P450依赖或独立型)。需要持续检测种群进化代谢抗性,来调整抗性管理策略,更好防治这些杂草。
3 SAH的感知、运输和代谢
3.1 生长素类除草剂的潜在靶标位点抗性机制:生长素受体
对于SAH,抗性候选机制可能包括靶标位点的改变(生长素受体或特异性生长素转运体)和如代谢SAH的其他转运体和酶的非靶点机制。除草剂抗性的1个主要来源为靶标位点抗性。靶标位点抗性频繁产生于除草剂结合的蛋白质活性位点的突变,使蛋白质对一些除草剂不敏感。田间应用SAH产生抗性还没有被追溯到核心信号传导靶标位点。然而,许多这些蛋白的突变可对SAH产生抗性,如对SAH有耐性的拟南芥突变筛选确定大多数生长素信号传导和运输蛋白。
科学家已确定许多生长素受体蛋白,而受体TIR1为典范。对于拟南芥,TIR1为6个受体组成的家族的一部分,其他 5个为生长素 F-盒蛋白(AFB1-5)。此家族由3对受体组成:TIR1和AFB1,AFB2和 AFB3,AFB4和 AFB5。所有受体间具有很高程度的序列相似性,很高程度的功能冗余。其中任一基因功能突变所致植物基因型与野生型相比几乎没有变化,只有叠加突变才使植物的生长和发育受到影响。
TIR1受体结构通过结晶化测定。通过研究TIR1结合生长素和共受体蛋白 AUX/IAA的特性了解TIR1的作用机制(图 6)。通过筛选大量的生长素和类生长素分子已开发了药效图,这些图能给出每个受体的活性配体的详细特性。
TIR1和AFB1-5生长素受体的序列几乎不变,在植物界有高度的序列保守性,特别是结合位点的氨基酸。这表明1处突变会使植物的适合度下降,任何与野生型的偏离都是不利的。如果这是正确的,那么靶标位点抗性不可能产生。受体家族的过多就会使植物系统灵活,因此一个受体效力的减少可被其他补偿。适合度的降低只存在于对内源性生长素IAA敏感的情况。因此,了解是否TIR1家族存在初期阶段抗性和是否受体家族不同成员对不同 SAH有不同选择性很重要。有证据表明AFB5和TIR1对不同SAH的选择性不同,AFB5受体是吡啶甲酸除草剂的主要作用位点,也有证据表明对氨氯吡啶酸不敏感的拟南芥AFB5突变品系对2,4-滴或IAA的敏感性没变。因此,杂草种群的AFB5同源物可能发生突变,从而对吡啶甲酸除草剂产生抗性,而对内源生长素的敏感性或植物的适合度没有改变。吡啶甲酸除草剂管理需要说明和阻止此种情况的出现。
共受体 AUX/IAA蛋白应被考虑,因为一旦生长素在识别口袋里,共受体蛋白就结合在生长素的顶部形成共受体复合体(图6)。不同AUX/IAA与不同浓度生长素结合,共受体的突变会造成对SAH产生抗性这是可能的。基于生长素受体和共受体的靶标位点抗性仍没有被确定,但可能存在。可能由于受体家族的过剩,不同骨架的SAH会保持活性。
3.2 生长素类除草剂的潜在靶标位点抗性机制:生长素运输蛋白
运输蛋白能够赋予靶标位点和非靶标位点抗性(图7)。植物细胞主要是通过AUXin抗性1(AUX1)载体选择性吸收生长素。最初从对 2,4-滴抗性的植物体内分离到AUX1突变体,已知这个靶标位点易发生变化。AUX1结构仍没有被描述,但可用烟草细胞培养进行放射性标记的 2,4-滴累积生测来测定AUX1的活性。已研究了多样的类生长素结构,预期可得到AUX1选择性的药效图。数个SAH骨架不是AUX1的底物,但化合物仍与SAH一样有活性。所以,SAH抗性好像不可能由AUX1突变产生或质膜中AUX1丰度的降低所致。

图6 与2,4-滴结合的生长素受体TIR1的结构

图7 生长素运输蛋白的排列
选择性生长素外流产生了极性生长素运输,使植物细胞产生极性并维持。有2个重要的外流载体蛋白家族 PIN-FORMED(PIN)和 ABCB蛋白。PIN具有植物特异性,ABCB是引起药物抗性产生的ATP驱动转运体家族的一部分。假如PIN和ABCB是复杂的膜结合蛋白,那么目前其结构未知。此外,没有适于研究它们药理学的广泛使用的转运生测。已知有少许选择性运输抑制剂,其中萘基酞基氨酸已被登记为除草剂。
更具有挑战性的是在已有生化知识基础上评估外流蛋白作为抗性位点的潜力。外流载体的上调可能促进底物生长素从细胞中排出,通过清除除草剂产生抗性。在考虑内源性生长素浓度和梯度时,PIN起主要作用。对PIN对不同SAH的选择性知之甚少,但已知PIN的活性为自我增强型,因此增加生长素剂量可能补充或减少质膜中 PIN的损失而增加活性。高剂量SAH能促进PIN补充,但它们如何影响流出和抗性将取决于PIN蛋白对除草剂的选择性。生长素输送机制和PIN蛋白的药理选择性的更多知识有助于解释PIN在SAH作用中的角色。
ABCB运输蛋白在非植物真核表达系统中被很好地表征,目前正在不断地了解植物中ABCB运输蛋白的重要性。植物ABCB的初步同源模型已被提出,但还不足以精炼用于了解底物的选择性。植物ABCB与生长素外流有特异性相关性,与依赖于它们正确的功能的物理反应有关。此家族具有许多小分子底物和质膜的ABCB丰度的变化导致对小分子药物的抗性。更好地了解此家族植物运输蛋白关系到避免 SAH抗性的进化。对 2,4-滴抗性因情形与ABCB转运体活性有关,虽然导致细胞内累积量降低的受损的长距离运输和没有增加的细胞外排好像是抗性的基础。
3.3 生长素类除草剂的潜在非靶标位点抗性机制:生长素的代谢
农药的非靶标位点抗性与酶把农药代谢为无害物质有关。使内源生长素IAA失活的主要途径为结合。结合IAA的酶可能被认为是使合成生长素失活的候选物和抗性的潜在原因。植物体内IAA结合有3种主要途径:通过GH3蛋白,生长素与氨基酸结合;通过葡萄糖基转移酶与糖结合;通过 GST3与谷胱甘肽结合。已确定与IAA和水杨酸结合的GH3蛋白结构。原则上,结合酶可能是抗性产生的原因,但任何一个酶识别所有不同 SAH骨架为底物是不可能的。
通常细胞色素P450酶对小分子的氧化,可能是除草剂和包括杀虫剂在内的其他农药解毒的一个机制。植物对二氯喹啉酸抗性与这些酶有关。主要催化内源生长素IAA的途径被确定为是IAA氧化酶,而不是细胞色素P450。是否此酶也将以SAH为底物还未知。虽然内源性IAA氧化酶可能是抗性产生原因,但对多种SAH的抗性是不可能的。氧化已成为最近开发耐麦草畏和 2,4-滴作物的机制。在这些情况下,细菌来源的2种不同的氧化酶被引入赋予抗性。
4 SAH发现的最近创新
在20世纪90年代晚期氯氨吡啶酸的发现促进了陶氏益农对吡啶羧酸SAH结构-活性关系的研究。这导致发现1类新颖SAH,即吡啶羧酸的衍生物芳基吡啶甲酸类。SAH的酸部分功能对其在植物韧皮部中的移动和通过离子捕获在细胞中的累积重要。对于吡啶羧酸除草剂二氯吡啶酸、氨氯吡啶酸、氯氨吡啶酸和芳基吡啶甲酸类除草剂[氟氯吡啶酯(ArylexTMActive)和氯氟吡啶酯(RinskorTMActive)],羧酸为与TIR1和AFB1-5受体进行主要结合的功能团。芳基吡啶甲酸被制剂化为酯,这些酯在植物体内可能迅速水解为游离羧酸。
芳基吡啶甲酸具有独特的除草活性,与吡啶羧酸除草剂相比相对低的使用剂量(相当于30 g酸/hm2或更少)防效增加。类似物DAS402除草活性显著高于氯氨吡啶酸(图8),但其土壤半衰期太长(大于240 d),不能满足预期商业用途的法规要求。在DAS402结构中加入可代谢的部分,可增加其在土壤中的降解。这导致发现了 4-氨基-3-氯-6-(4-氯-2-氟-3-甲氧基苯基)-2-吡啶甲酸甲酯(氟氯吡啶酯)(图8)。氟氯吡啶酯是活性高的广谱SAH,对小麦和大麦高度安全,土壤半衰期为10-30 d,比DAS402的半衰期少很多。在2015年被首次商业化用于谷物。
在氟氯吡啶酯的开发过程中, 吡啶甲酸骨架的数个重要特征被确定:分子的2-羧酸和4-氨基功能团对高活性和广谱性重要;3-氯功能团进一步增强了除草活性。对氟氯吡啶酯的吡啶甲酸部分5-位的SAR的研究发现了另一有商业用途的芳基吡啶甲酸类除草剂。氟氯吡啶酯的5-氟类似物不但具有高活性和广谱性,而且对水稻有优异的选择性。其相对应的羧酸[4-氨基-3-氯-6-(4-氯-2-氟-3-甲氧基苯基)-5-氟-2-吡啶甲酸]的土壤半衰期为 10~30 d。进一步优化开发了氯氟吡啶酯,用于水稻和其他作物田防治一年生禾本科杂草稗草和一些莎草科杂草(莎草属)。在2017年首次获批登记。
芳基吡啶甲酸除草剂的发现有力地表明有许多机会来开发新的SAH。芳基吡啶甲酸活性的多样性使SAH更广泛地用于杂草管理市场,如谷物、水稻、牧场和道路用地。
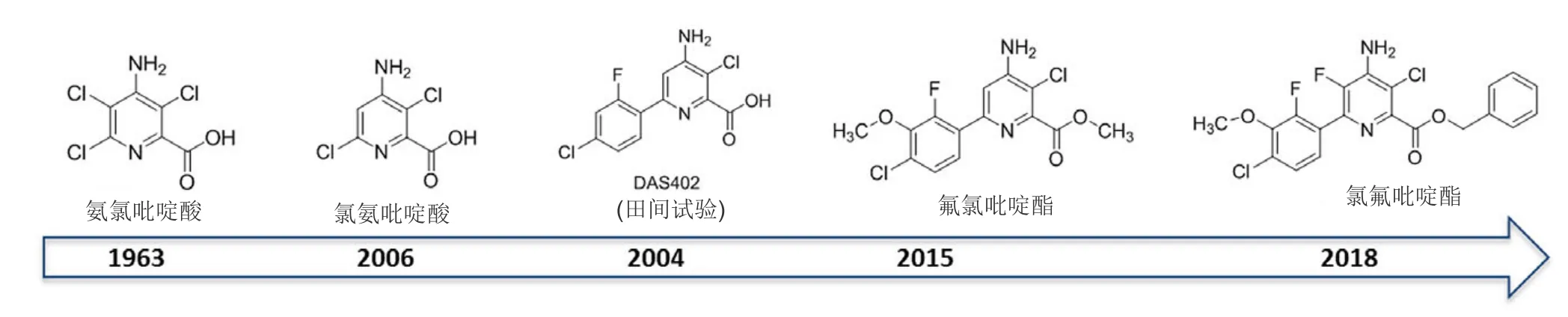
图8 在相隔43年中发现了氨氯吡啶酸和氯氨吡啶酸
5 管理耐SAH作物的抗性
已开发了新的除草剂制剂用于陶氏益农最近开发的耐 2,4-滴作物和孟山都开发的耐麦草畏作物。Enlist DuoTM(2,4-滴-胆碱+草甘膦)和 Enlist OneTM(2,4-滴-胆碱)除草剂被开发用于EnlistTM杂草防治系统和采用VaporGrip®技术生产的Xtendimax(麦草畏)被用于Roundup Ready®Xtend作物系统。陶氏益农和孟山都已开发了管理项目以确保这些耐除草剂作物系统能被长期持续地使用,为把其整合入除草剂抗性管理项目提供指导。此外,美国环保局(USEPA)正在制定规章制度,强调需要种植者采用最好的除草剂抗性管理措施以进一步持续使用这些作物系统。
随着抗SAH作物的引入,从监管和管理角度广泛考虑了除草剂的抗性管理。这些工具将有助于进行杂草管理项目和防治一些抗除草剂杂草品种。广泛采用的延缓杂草种群除草剂抗性选择和管理现有抗性杂草种群的最好的管理措施形成管理这些技术的基础。这些最好的管理措施是美国USEPA规定的登记条件的重点。
从20世纪50年代到2001年,在美国,除草剂登记者必须满足有关除草剂抗性管理的少许要求。在 2001年后登记者被要求用不利影响报告程序(Adverse Effects Reporting process)向USEPA报告已确认的抗除草剂杂草品种。此外,USEPA表明登记者可能自发地在除草剂产品标签上表明作用机制,标明除草剂抗性和阻止或延缓除草剂抗性进化的最好管理措施。在2014年,USEPA首次发布了Enlist Duo除草剂登记条件,要求要具有除草剂抗性管理计划。在2016年,发布了Xtendimax (采用Vaporgrip技术生产)除草剂的相似批准条件。这些 SAH杂草管理系统的 USEPA除草剂抗性管理计划由田间检测和补救,培训和信息,评估,最好的管理措施和报告组成。
需要利益相关者间加强合作来开发和实施有效的除草剂管理项目,限制田间抗性进化,确保除草剂能被长期持续地使用。同时除草剂开发者和登记者、农民、管理者、推广者、专家、作物顾问、经销商和零售商也要进行技术管理。行业鼓励杂草管理相关利益方继续促进把除草剂作为综合的杂草管理方法的一部分,通过延迟田间除草剂抗性进化使这些有价值的工具被继续使用。
6 总 结
本文目的是介绍SAH的植物抗性知识,为未来开发SAH产品带来更好的机会。SAH药理学独特、复杂。其复杂的作用机制,对农业生产有益的影响和持久性是其用于杂草管理体系的自白书。
SAH在商业使用 70多年后可被继续应用,部分原因为它们一般不被作为杂草防除的唯一工具。最近对SAH的作用机制和作用位点的观点,有过剩的生长素受体、生长素反应型基因的改变和多个生化途径的动态平衡的中断,这说明不能以简单假说解释许多田间抗性生物型产生的原因。这也说明靶标位点抗性发展慢的原因,以及可能和适合度代价有关。最近采用最新分子和生化工具进行的新抗性杂草生物型的调查为未来抗性风险(特别是非靶标位点抗性机制)提供了信息和为新 SAH提供了潜在的延缓风险的策略。
目前也难以证实简单的作用位点改变导致对SAH亲和性下降而产生抗性。代谢增加和植物吸收降低的假设常不足以解释生物型对SAH的反应。在对化学、蛋白质和基因调节间相互作用的了解下和应用改进的分析工具,现在了解了运输的降低可能为 SAH抗性产生的潜在原因,增加的代谢(例如虞美人)可能或不可能(例如刺莴苣和野萝卜)与 SAH抗性有关。对这些机制每一种影响的了解已证明是困难的,因为在复杂生长素反应途径中存在多个和冗余的变量。
在笔者收集的观点中,由于靶标位点(结合蛋白,TIR1/AFB1-5))改变导致的抗生物型的风险相对低。笔者也认为生长素感知和反应途径的复杂相互作用通过靶标蛋白的过度表达(抗草甘膦地肤和长芒苋)形成的抗性可能性也低。由于 SAH的使用继续增加,抗性选择风险将也增加。笔者认为螯合作用和/或抗性运输机制的改变(例如 ABCB转运体,其他)可能发展和扩展。需要重点关注导致对其他除草剂作用机制共进化抗性的对 SAH运输改变的性状的选择,这在多抗药细菌已发现。SAH的差异代谢也已在一些杂草生物型发现,了解这些抗性机制对开发和实施更有效抗性管理策略重要。
通过增加知识和分析能力可能获得这些见识。但是,对细胞的药理学和植物体内长距离运输机制了解的少。杂草科学和联合学科为更好地了解这些机制提供了机会。在使用70多年后,具有独特的杂草防治谱和不同化学特性新 SAH继续被发现的事实支持此观点,即未来有很多机会来进一步对SAH进行创新。
另一经验为整个杂草防除专家群体包括行业科学家、学术界、管理机构和农民需要意识到和充分支持实施最好的除草剂抗性管理措施。“多样性”是维持有效长期杂草管理的最主要的原因。例如合用不同作用机制的土壤残留除草剂和芽后除草剂以及非化学杂草管理方法(即覆盖作物、策略耕作、作物轮作和收获时杂草种子防治措施)将可能会使杂草管理工具切实可行,能使现代农业蓬勃发展,为后代提供安全的粮食。
