垂直切根对当年羊草构件及繁殖资源分配的影响
2018-12-27陈积山孙泽微
王 欢,邓 波,陈积山,孙泽微
(1.吉林农业大学动物科技学院,吉林 长春 130118; 2.中国农业大学动物科技学院草业系,北京 100193;3.黑龙江省农业科学院草业研究所,黑龙江 哈尔滨 150086)
羊草(Leymuschinensis),又名碱草,为多年生根茎型禾本科牧草,分布在欧亚大陆的东部地区,是我国北方重要的牧草资源,对改善北方草原生态环境有重要意义。羊草根茎主要分布于0-15 cm土层中,在地下横走形成网状,多年生羊草根茎致密,会降低土壤理化性质。松土改良通过改变土壤的理化性质和刺激植物根系两方面使原生植被恢复[1]。切根是草地松土改良的方式之一,依据羊草根茎自我繁育的生物学特性,利用垂直刀具切断羊草的地下横走根茎,可促进羊草根茎复壮与自我繁殖[2-3],增加群落盖度、密度、生物量和竞争能力[4-7],降低草地毒杂草比例[8]和土壤容重[9-10],改变土壤理化性质[8,11],加速退化草地群落的恢复演替。
植物的繁殖分配是指植物一年所同化的资源中用于生殖的比例[12],也是植物种群生殖器官(穗、花、果实)耗用或积累的干物质、营养元素或能量,在整个种群生活史中总耗用和积累总量中所占的比例[13],这种分配比例的变化可以反映植物的繁殖对策。人们对其的研究常采用将植物分为与其结构、功能相关植物构件(叶、茎、穗、根等)的干重、营养元素和热值分析的方法。繁殖分配不仅是植物种群的重要选择适应特征,而且与环境条件存在密切关系[14]。Sakai[15]、Gardner和Mangel[16]认为,有利和多变的生境会促进克隆植物的有性繁殖,无干扰的环境中,营养繁殖在选择上更占优势。也有研究表明,在生长限制的条件下,克隆植物的分配比例往往更倾向于营养繁殖并较少地进行有性繁殖[17-19]。这可能是植物个体间的差异和种群繁殖对策受资源水平的影响,因此,有必要对了解植物的繁殖分配的因素进行更多的研究工作。
羊草具有丰富的遗传多样性,是研究克隆植物适应性进化和种群生态学的理想植物[20]。目前对于切根的研究多见于不同深度与间距对羊草种群特征的影响,对于不同切根时期与间距互作条件下,羊草生物量、营养元素及能量等资源的繁殖分配策略鲜见报道。为此,在羊草不同发育时期进行不同间距的切根处理,对羊草繁殖分配特征进行研究,为探明羊草适应和抵抗逆境采取的生活史对策、确定适合羊草草地物理改良的方法、建立退化草原改良体系和规范提供理论基础和技术支撑。
1 材料与方法
1.1 研究区概况
试验样地为天然羊草草地,位于松嫩平原西部,黑龙江省绥化市境内的国家牧草产业技术体系绥化综合试验站内(46°32′17″ N, 125°28′24″ E)。该地区平均海拔160 m,属于温带大陆性气候,年降水量351 mm,降水集中在6-8月,冬夏季风更替明显。年均日照时数2 713 h,年均气温4 ℃,极端最高气温与最低气温分别为40 ℃、-33 ℃,年均初霜期为9月23日,年均终霜期为5月15日,全年结冻期为183 d。土壤类型为碱化草甸土,土壤有机质、全氮、全磷含量分别为2.981、0.301、0.416 mg·kg-1,土壤全盐量变化范围为0.157%~0.318%,土壤呈现轻、中、重三级盐碱化,pH 8.12~10.08。试验区优势种为羊草、星星草(Puccinelliatenuiflora),伴生种主要有虎尾草(Chlorisuirgara)、冷蒿(Artemisiafrigida)、灰绿藜(Chenopodiumglaucum)、蒲公英(Herbataraxaci)、委陵菜(Potentillachinensis)等。试验区天然草地牧草5月中上旬返青,7月至8月上旬为牧草的旺盛生长期,9月初进入枯黄期。试验当年,草地在5月初返青。
1.2 试验设计
2017年5月12日,选择植被生长均匀、盖度均一、地势相对平坦的天然羊草草地。样地上进行定桩、拉线划方、实施切根。切根方式为“井”字形切根,切根深度为12 cm。自制切根刀具(长25 cm,高15 cm,厚0.5 mm)。试验设定切根时期和切根间距两个因素,不切根为对照,其中切根时期分别为返青期(R)、孕穗期(B)和抽穗初期(H),切根间距(宽度)为10、20和30 cm,即L10、L20、L30。两因素组合共计9个处理(表1),1个对照,每个处理3次重复,小区面积为3 m×5 m,共计30个小区,小区周围设置1 m隔离带,小区布设采用随机区组设计。
表1 切根处理设置Table 1 Root cutting treatments
1.3 测定指标及方法
构件收集:于2017年8月,在每个小区内随机取6个长×宽为25 cm×25 cm的样方内将羊草齐地面刈割,并将样方切成 25 cm×25 cm×25 cm的土柱,把土柱放入纱网袋带回实验室,浸泡并冲洗干净后风干,将样品按叶(叶与叶鞘)、茎、穗(穗与穗柄)和根茎(根与根茎)等构件分开。
生物量分配:将羊草构件分别装入纸袋内,在80 ℃下烘干至恒重后称重。叶、茎、穗之和为地上生物总量,种群总生物量包括地上生物总量与根茎生物量,计算每平方米构件生物量及构件生物量占羊草地上与地下总生物量的百分比。
营养元素分配:将烘干样品粉碎,采用全自动凯氏定氮仪(2300 Kjeltec Analyzer Unit,FOSS,Sweden)测定各构件的全氮(N)含量,钼锑抗比色法(UH5300)测定各构件的全磷(P)含量。羊草各构件N、P元素总量为各构件生物量与N、P含量的乘积,羊草各构件N、P元素的分配为各构件N、P总量占羊草N、P总量的百分比。
能量分配:称取烘干粉碎后的构件样品,用ZDHW-6微机全自动量热仪检测羊草单位生物量的热能值。各构件能量累积值为该构件的热能值×生物量;构件能量分配比为该构件能量值占羊草总能量的百分比。
1.4 数据处理
数据采用Excel 2010 制作图表,用SPSS 19.0统计软件中的一般线性模型对切根时期和切根间距进行两因素方差分析(two-way ANOVA),以说明其交互作用。对各切根处理进行单因素方差分析(one-way ANOVA),差异显著性(P<0.05)用Duncan法进行多重比较。
2 结果与分析
2.1 切根对羊草构件生物量及其分配的影响
双因素方差分析得出,切根时期对羊草穗与根茎生物量影响均达到极显著水平(P<0.01)(表2),同一切根间距下,穗生物量随着切根时期延后而增加,根茎生物量在返青期切根明显增加,切根时期对羊草叶与茎生物量无显著影响(P>0.05)(表3);切根间距对羊草叶、茎、穗生物量影响均达到极显著水平(P<0.01)。同一切根时期,叶与穗生物量均随切根间距增加而增大;切根时期与间距的交互作用对茎生物量影响显著(P<0.05),与对照相比RL10与HL20处理茎生物量显著下降。RL10处理地上生物总量与对照相比显著降低;切根间距30 cm时,地上生物总量随着切根时期延后而增加相同切根时期,地上生物总量随切根间距增加而增大。羊草构件生物量大小为根茎>叶>茎>穗,但BL30与HL30切根处理下叶>根茎>茎>穗生物量,整体来看,30 cm切根间距提高了羊草地上构件生物量,返青期切根提高了根茎生物量。
切根间距对羊草构件生物量分配比例影响明显(图1)。返青期切根,穗的生物量繁殖分配比例降低,根茎生物量分配增加,其中RL10处理根茎生物量较对照提高8.62个百分点,且随着切根间距增大茎、穗生物量分配也增长,根茎分配降低;孕穗期与抽穗初期切根,根茎与穗的生物量分配降低,叶的分配比例增加,且随着切根间距增加叶、穗生物量分配也增加,根茎分配比例降低,其中抽穗初期切根间距30cm处理,羊草穗的生物量繁殖分配比例为2.32%,较对照高0.17个百分点(图1)。

表2 切根时期与间距交互作用下羊草各构件生物量的方差分析(F)Table 2 Variance analysis of Leymus chinensis biomass in root cutting treatments (F)
*,**分别表示在0.05和0.01水平差异显著。下同。
* and ** indicate significant difference at 0.05 and 0.01 level,respectively; similarly for the following tables.
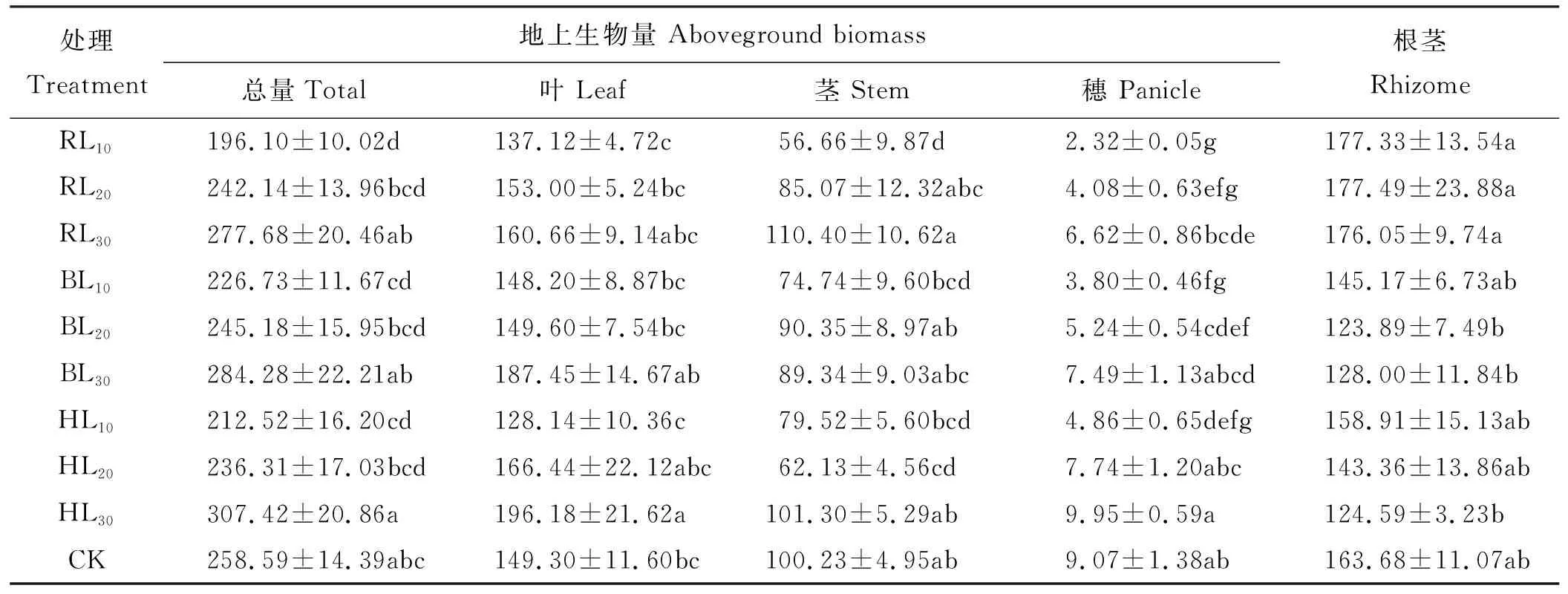
表3 切根对羊草构件生物量的影响Table 3 Effect of root cutting on Leymus chinensis biomass g·m-2
RL10,RL20,RL30,BL10,BL20,BL30,HL10,HL20,HL30,同表1,CK为不切根对照。同列不同小写字母表示在0.05水平上差异显著,下同。
RL10, RL20, RL30, BL10, BL20, BL30, HL10, HL20, HL30, were the same at Table 1; CK, no root cutting. Different lowercase letters within the same column indicate significant difference at the 0.05 level; similarly for the following tables.
相同构件不同小写字母表示在同一切根时期不同切根间距差异显著(P<0.05)。下同。
Different lowercase letters within the sameLeymuschinensiscomponent indicate significant difference between different cutting distance at the 0.05 level; similarly for the following figures.
2.2 切根对羊草构件营养元素含量及分配的影响
双因素方差分析得出,不同切根时期对羊草根茎全氮含量影响达到极显著水平(P<0.01)(表4),对穗全氮含量有显著影响(P<0.05);不同切根间距对茎、穗全氮含量影响均达到极显著水平(P<0.01),对叶全氮含量有显著影响(P<0.05);但两者的交互作用仅对根茎全氮含量影响达极显著水平(P<0.01),对其他构件全氮含量无显著影响。所有处理对全磷含量无显著影响(P>0.05)。切根明显提高了羊草叶、茎、穗的全氮含量)(表5),其中RL10处理对羊草叶、茎、穗的全氮含量影响显著(P<0.05);而RL30处理显著提高了羊草根茎的全氮含量, BL30处理显著降低了根茎的全氮含量,其他处理对羊草根茎的全氮含量影响不显著(P>0.05)。
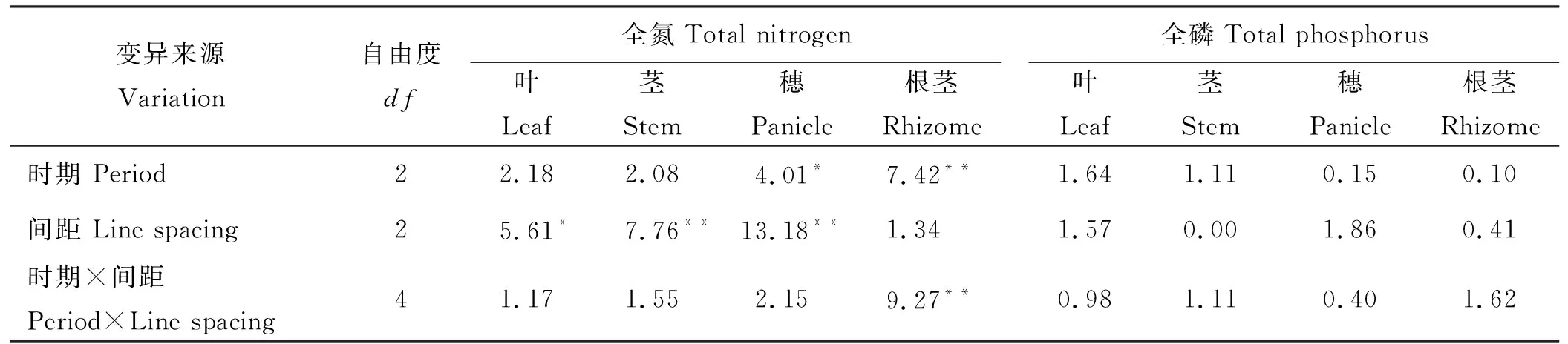
表4 切根时期与间距交互作用下羊草各构件全氮与全磷含量的方差分析(F)Table 4 Variance analysis of total N and P contents in Leymus chinensis root cutting treatments(F)
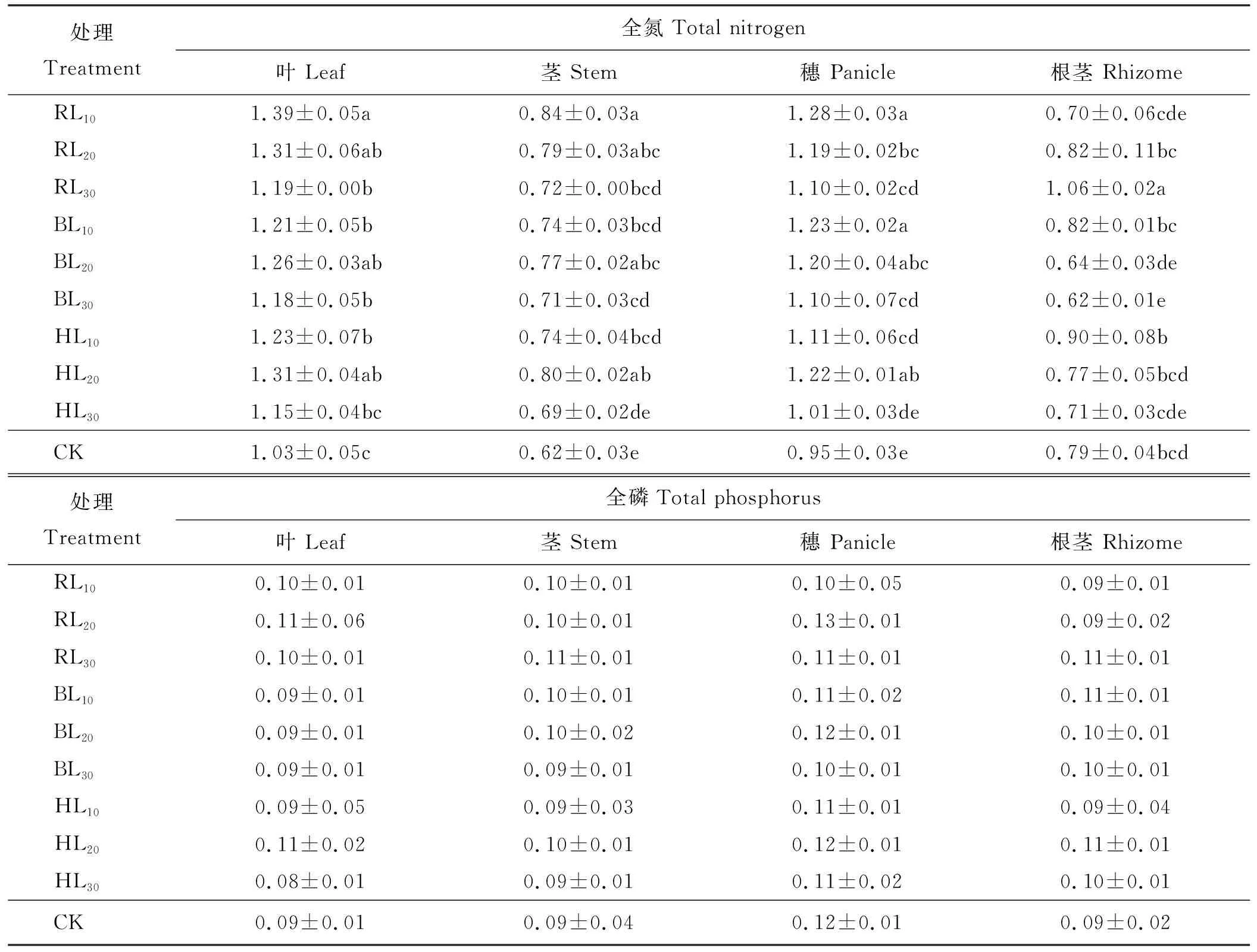
表5 切根对羊草构件全氮与全磷含量的影响Table 5 Effect of root cutting on total N and P contents in different Leymus chinensis components %
切根对羊草叶与根茎的全氮分配影响显著,对羊草茎的全磷分配影响显著(P<0.05)(表6)。切根有利于羊草叶的全氮分配,仅RL30处理低于对照5.26%; BL20、BL30、HL20、HL30处理显著降低了根茎的全氮分配; RL10与HL20处理下茎的全磷分配比例较对照显著降低。切根对羊草穗的全氮与全磷分配影响显著(P<0.05),相同切根时期,穗的全氮与全磷分配比例均随切根间距增加而增加;相同切根间距,穗的全氮与全磷分配比例随切根时期的推移而增加。
2.3 切根对羊草构件能量累积及分配的影响
双因素方差分析得出,不同切根时期对羊草穗和根茎能量累积影响达到极显著水平(P<0.01)(表7),不同切根间距对叶、茎、穗能量累积影响均达到极显著水平(P<0.01),但两者的交互作用仅对茎的能量累积有显著影响(P<0.05)。
由于切根对羊草构件生物量有显著影响,致使切根对羊草叶、茎、穗的能量累积也具有显著影响(P<0.05)(表8)。同一切根时期,叶、穗的能量累积随切根间距的增加而增大;同一切根间距,穗的能量累积量随切根时期的推移而增加;RL30与HL30处理提高了茎的能量累积量;相同切根间距,不同切根时期根茎的能量累积顺序为返青期>抽穗初期>孕穗期。整体来看,HL30处理有利于地上构件能量累积,RL10处理利于根茎能量累积。
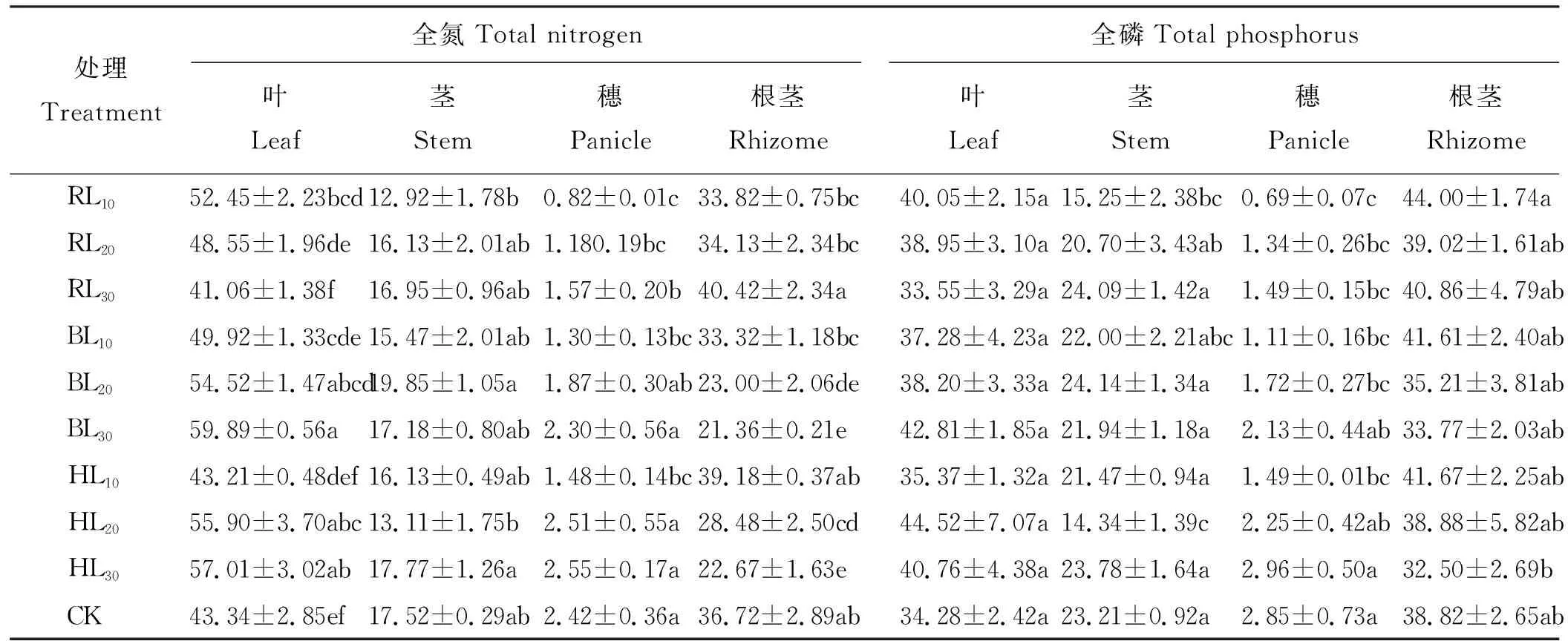
表6 切根对羊草构件全氮与全磷分配的影响Table 6 Effect of root cutting on total N and P distribution in Leymus chinensis components %

表7 切根时期与切根间距交互作用下羊草各构件能量累积的方差分析(F)Table 7 Variance analysis of energy accumulation in Leymus chinensis root cutting treatments(F)
切根显著影响羊草能量在构件间的分配,即叶、茎、穗、根茎的累积能量在不同切根处理下所占比例有差异。孕穗期与抽穗初期切根对羊草构件能量分配比例影响显著(P<0.05),而返青期切根对构件能量分配无显著影响(P>0.05)。同一切根时期,叶的能量分配随间距增加呈升高趋势,根茎能量分配随切根间距增加而减小,其中RL10处理下根茎的能量分配比例较对照提高21%。
在孕穗期与抽穗初期切根,对羊草穗的能量繁殖分配比例影响明显(图2)。羊草穗能量繁殖分配随着切根间距增加而增大;切根间距10 cm,能量繁殖分配表现为孕穗期<返青期<抽穗初期;切根间距10 cm的处理降低了羊草穗的能量繁殖分配;而HL30处理的能量繁殖分配比例为2.49%,提高了羊草穗的能量繁殖分配。

表8 切根对羊草构件能量累积的影响Table 8 Effect of root cutting on the energy accumulation of Leymus chinensis components kJ·m-2
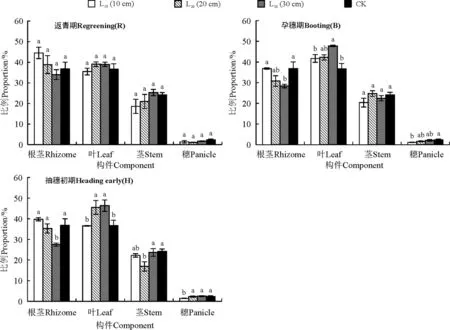
图2 切根对羊草能量繁殖分配的影响Fig. 2 Effect of vertically cutting root on energy distribution in Leymus chinensis
3 讨论
在克隆植物完成生活史过程中,会对满足自身有性繁殖或无性繁殖的所需资源进行权衡分配,这种繁殖分配不仅是繁殖对策的重要内容,也是种群延续和不断扩张的关键[21]。虽然植物的繁殖对策很大程度上是由遗传决定,但不同的环境条件下同一物种繁殖分配也有着很大的可调节性,与海拔、水分、气候条件、营养水平、群落特征和不同生境等因素存在密切关系[22-28]。
有性繁殖和营养繁殖是植物的两种繁殖方式,当受到环境干扰时,植物通过调节资源配置提高适合度应对环境变化。本研究得出,切根作用下,各构件(叶、茎、穗、根茎)营养元素全氮、全磷、能量的分配比例与种群生物量分配比例变化基本一致,这与王仁忠等[33]得出的羊草构件生物量分配和能量分配是等价的观点相吻合。同时,本研究表明,羊草返青期切根和间距10 cm切根,根茎的资源分配达到最高水平,穗的繁殖分配程度最低,增加了羊草无性繁殖比例,同时抑制了有性繁殖,这与杜利霞等[29]、Van Groenendael和Habekotte[30]的结果相符,即资源限制时,生殖格局由有性繁殖和无性繁殖并存转向单一的无性繁殖转变。而与Loehle[31]的结果不一致,原因可能是返青期羊草植株较弱,同时10 cm切根对羊草根茎损伤严重,不能及时向地上部分供应养分,进而限制了羊草根茎资源的运输与分配。而抽穗初期切根间距30 cm最大程度的提高了羊草的繁殖分配,促进了羊草有性繁殖,表明适宜的生存环境更利于羊草有性生殖,这与Sakai[15]、Gardner 和Mangel[16]的结论相一致,但与Herben[32]的结果(有利的生存环境会促进克隆植物的无性繁殖)不一致。造成这些矛盾的主要原因可能是,克隆植物的有性繁殖和无性繁殖权衡分配不仅受植物个体资源水平的影响,也受植物种群密度、高度等种群特征和种间竞争及环境因素的共同影响。
切根处理后叶、茎、穗、根茎等构件的资源分配是羊草对所处环境条件做出的适应性进化特征,返青期切根提高了羊草根茎生物量、促进了根茎的复壮更新,30 cm间距切根可以增加羊草地上生物量,切根后羊草各地上构件的全氮含量增加,这与“切根可以提高羊草的粗蛋白含量”[3]结果一致。整体来看,同一切根间距处理下,不同切根时期羊草繁殖分配表现为抽穗初期>孕穗期>返青期,可能原因是随着羊草生长发育,自身抗干扰能力增强。而同一切根时期,羊草繁殖分配的表现为切根间距30>20>10 cm,说明10 cm切根间距是不适合羊草繁殖分配的切根处理。研究还发现,羊草的繁殖分配不足5%,而羊草草地改良的限制因素主要是种子产量低,因此羊草繁殖分配低也可能是羊草种群种子低产的重要原因之一。
4 结论
切根显著影响了羊草穗的繁殖分配和叶、茎、根茎的资源(生物量、全N、全P、能量)分配,抽穗初期间距30 cm切根对繁殖分配影响达到最佳切根效果,有利于羊草的有性繁殖。而返青期间距10 cm切根提高了根茎的资源分配比例,促进了羊草的无性繁殖。总体来看,切根干扰对羊草的资源分配影响显著,适宜的切根处理可以改变羊草的繁殖分配策略。
