设施菜地种植年限对土壤理化性质和生物学特征的影响
2018-12-20王学霞陈延华王甲辰
王学霞,陈延华,王甲辰,左 强,肖 强
(北京市农林科学院植物营养与资源研究所,北京100097)
设施菜地栽培因其集约化程度高、产能高效、农业环境适应能力强等特点,可运用生物、工程与环境技术,使农业生产不再受限于气候、季节等因素,有效地提高了农业资源综合利用效益和水平[1],在我国发展迅速,在蔬菜反季节和跨地区种植中起着重要作用,成为我国农业经济的重要来源之一[2–3]。随着北京都市型现代农业发展要求,以及农业产业结构调整和供给侧改革的需求,设施菜地在北京农业中占据比重逐年增大[3–4]。
设施菜地土壤处于长期的高集约化栽培、高复种指数、高肥料施用量的生产状态,菜地土壤理化性状和生物学特性发生了很大变化[2,4–6]。随着种植年限的增加,土壤次生盐渍化严重,土壤速效氮、磷、钾等营养离子在土壤表层大量积累,尤其是NO3–-N不断累积[7–8],稳性团聚体比率逐渐降低[9],影响了作物根系的生长,并破坏作物对养分的均衡吸收[10–11]。土壤次生盐渍化还导致土壤动物和微生物种群数量改变、功能多样性降低、群落结构失衡,土壤酶活性改变,土壤周转氮库减少,不仅直接危害作物的正常生长,也造成土壤质量的快速退化[12–13]。
土壤生物是生态系统的重要组成部分,作为土壤分解系统的主要驱动因子,能有效地评价土壤的活力和健康状况[14–15]。土壤微生物和土壤动物常年生活在土壤中,对土壤环境的变化敏感迅速,对种植年限带来的变化也会有所反应[12,16]。近年来对设施菜地土壤微生物的研究发现,随着种植年限增加,微生物数量或增加[17]、降低或者变化不明显[18],而土壤微生物群落结构变化明显,尤其是与氮循环相关的功能群微生物[19]。土壤线虫群落结构可表征土壤的健康水平,近年来的研究发现,随着种植年限增加,土壤线虫密度呈现先增加后减少的趋势,植物寄生性线虫数量增加,而食真菌线虫数量降低,线虫群落多样性和稳定性也呈现不同变化趋势[12,20–21]。
设施菜地不同种植年限土壤性质的变化可能导致土壤生物数量及群落结构的变化,因此,本项研究采集北京市延庆不同种植年限设施菜地土壤,系统研究比较了土壤理化性质、微生物数量、土壤线虫群落组成随种植年限的变化特点,为评价设施菜地土壤质量健康提供有效的生物指标,为设施菜地土壤可持续利用提供理论参考。
1 材料与方法
1.1 研究区概况
试验样地选取北京延庆旧县镇 (N 40°30′15″,E 116°05′45″),属大陆性季风气候。气候冬冷夏凉,年均降水量477.2 mm,年平均气温8℃,年日照2800 h。
1.2 试验设计
以露天菜地为对照 (CK),选择已种植蔬菜3年、5年、8年、12年的温室各 3个。每个种植年限设施菜地主要施用氮磷钾复合肥和氮肥,兼施少量有机肥,浇水方式为喷灌。设施菜地基肥施用氮磷钾复合肥1500~1725 kg/hm2(N∶P2O5∶K2O=15∶15∶15),追施氮肥量450~600 kg/hm2。露天菜田施用复合肥750~800 kg/hm2(N∶P2O5∶K2O=15∶15∶15),追施氮肥量300~375 kg/hm2。种植模式为果菜连作模式 (分春夏茬和秋冬茬),春夏茬主要种植番茄、黄瓜、茄子等果菜,秋冬茬主要种植油麦菜、菠菜等青菜。温室 (近5年) 在7月中旬到8中旬进行15天闷棚处理,种植过程中没有用药物杀线虫。
2017年5月25日取样。每个温室设5个小区(每个5 m × 5 m),每个小区按照“Z”字型取样,用直径3.0 cm土钻取土样,每个小区取10钻,土壤取样深度为0—30 cm充分混匀后作为一个样品,土壤样品分为3部分,一部分用于分析土壤理化性状,一部分4℃冰箱保存用于分离土壤线虫,一部分–70℃冰箱保存用于微生物数量分析。
1.3 测定方法
1.3.1 土壤理化性质测定 土壤容重采用环刀法测定;比重瓶法测定比重,计算土壤总孔隙度;电导率 (EC) 用电导仪测定法 (水土比5∶1);土壤pH测定利用酸度计 (水土比2.5∶1);、Cl–、K+、Na+、Ca2+、Mg2+养分离子采用滴定法测定;K+、Na+用火焰光度法测定[21]。土壤有机碳采用元素分析仪 (Vario EI,Elementar,German) 测定,和采用流动分析仪 (Auto Analyzer 3,SEAL,German) 检测分析。
1.3.2 线虫分离与鉴定 取 100 mL土壤用改进的贝曼浅盘法分离法分48 h。当分离出的线虫数量少于等于 100条时,鉴定所有的线虫种类,数量多于100条时,鉴定前100条,然后统计所有线虫数量。参照《中国土壤动物检索图鉴》[22]和《植物线虫分类学》[23]根据线虫的头部形态学特征和取食生境将土壤线虫划分到属。根据划分原则,将土壤线虫划分为以下 4 个营养类群[24–25]:食细菌类 (bacterivores,Ba)、食真菌类 (fungivores,Fu)、食植物线虫 (plantparasites,Pp)、捕食类/杂食类 (predators/omnivores,Om)。同时按照colonizer-persister(c-p),将线虫从c-p1到c-p5值分成4个群[26]。
1.3.3 土壤真菌和细菌数量分析 荧光定量PCR检测土壤微生物数量。取0.5 g土壤进行总DNA提取,每个样品3个重复 [Soil DNA Isolation Kit(MOBIO)] 作为样品DNA。以样品DNA作为扩增模版,用细菌16S rRNA基因的特异性引物 (F:GTGYCAGAM GCCGCGGTA;R:GGAC TACHVGGGTWTCTAAT),真菌18 s rRNA基因的特异性引物 (F:CGCTGCG TTCTTCATCG;R: TCCGTAGGTGAACCTGCGG)。在荧光定量PCR仪 (ABI 7500) 上进行扩增反应。荧光定量 PCR 扩增反应体系 (20 μL): 2 × Master Mix 10 μL(TaKaRa),10M 的 PCR 特异引物 F 0.5 μL, 10 μM 的PCR 特异引物 R 0.5 μL,加 ddH2O 7 μL,DNA 模版2.0 μL。反应程序: 95 ℃预变性 30 s; 40 个循环 95℃变性5 s,60℃退火30 s,72℃延伸30 s。为了建立PCR产物的熔解曲线,扩增反应结束后,按95℃,10 s;60℃,60 s;95℃,15 s;并从60℃缓慢加热到99℃(仪器自动进行-Ramp Rate为0.05℃/s),记录每个样品的CT值。将CT值代入标准曲线方程,计算出样品模版的初始基因拷贝数并最终换算出每克干土的细菌和真菌的数量。
1.4 计算公式
采用国内外学者广泛采用的Shannon-Weaver指数、Pielou均匀度指数研究土壤线虫的群落多样性[27]。
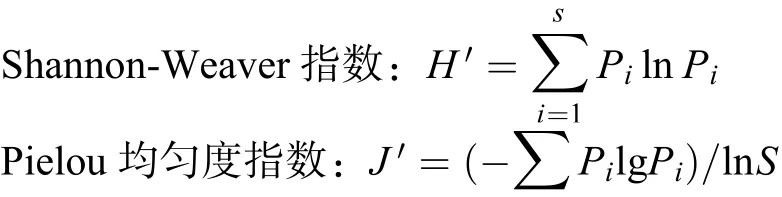
式中:Pi为土壤线虫群落第i类群个体数占总个体数的比例;S为土壤线虫群落中所有线虫类群数。
本文采用MI (Maturity index of free-living nematode)指数、PPI (Plant parasite index) 指数、NCR (Nematode channel ratio) 指数、WI (Wasilewska index) 指数表达各生境土壤线虫群落功能结构特征[27]。上述指数计算公式如下:


式中:v (i) 是在生态演替中对属于k-选择和r-选择的属分别赋予的c-p值;f (i) 是自由生活线虫属在线虫种群中所占的比例;f ′(i) 是植物寄生性线虫属在线虫种群中所占的比例;NBF为食细菌线虫数量;NFu为食真菌线虫数量;NPp是植物寄生线虫数量。
1.5 统计分析
所有原始数据经过对数转化保证方差齐性检验。分析过程运用SPSS 22.0 软件,运用单因素方差分析(one-way analysis of variance),差异显著性水平为0.05水平,文中数据均为平均数 ± 标准误。
2 结果与分析
2.1 土壤理化性质变化
表1表明,与对照 (CK) 相比,种植年限3、5、8、12年的土壤pH均有所降低,种植年限12年的土壤pH显著降低了0.92个单位 (P < 0.05)。土壤容重表现趋势与pH一致,已种植蔬菜12年的显著降低 (P < 0.05)。土壤总孔隙度、EC值、HCO3–、Cl–、SO42–、K+、Na+、Ca2+、Mg2+各种植年限均呈现为12 年 > 8 年 > 5 年 > 3 年 > CK,但差异达到显著的年限不同指标间有差异。与CK相比,土壤总孔隙度不同年限之间差异不显著;种植年限5、8、12年的土壤EC值分别显著增加了35.58%、68.46%、107.44%;种植年限12年的土壤HCO3–含量显著增加增加了37.04%;种植年限8年、12年的土壤Cl–含量分别增加了83.00%和169.71%;种植年限12年的SO42–含量显著增加29.71%;种植年限3、5、8、12年的K+含量分别增加了35.91%、135.04%、178.93%、267.04%;种植3、6、12年的Na+含量分别显著增加了18.97%、75.26%和 136.78%;种植年限 5、8、12年的 Ca2+含量分别显著增加了56.62%、101.22%和124.54%,土壤Mg2+含量分别显著增加了27.25%、29.49%和35.77% (P < 0.01)。土壤有机碳、NH4+-N和NO3–-N含量各种植年限均呈现 12 年 > 8 年 > 5 年 > 3 年 > CK。与对照 (CK) 相比,种植年限12年的土壤有机碳、NH4+-N含量分别显著增加了36.21%和29.51%;3、5、8、12年的NO3–-N含量分别显著增加了65.85 %、185.93%、302.85%和560.91%。
2.2 土壤线虫群落组成
图1显示,土壤线虫密度随种植年限发生了显著变化,表现为 12 年 > 5 年 > 8 年 > 3 年 > CK(P < 0.05),除种植年限5年与8年之间差异未达到显著水平,其余年份间均达到显著水平。种植年限3年、5年和8年土壤线虫密度增加量分别为16.74%、64.49%和58.22%,12年更是增加了93.19%。

表1 不同种植年限土壤理化性质 (均值 ± SE)Table 1 The physical and chemical properties of soils under different planting ages (mean ± SE)
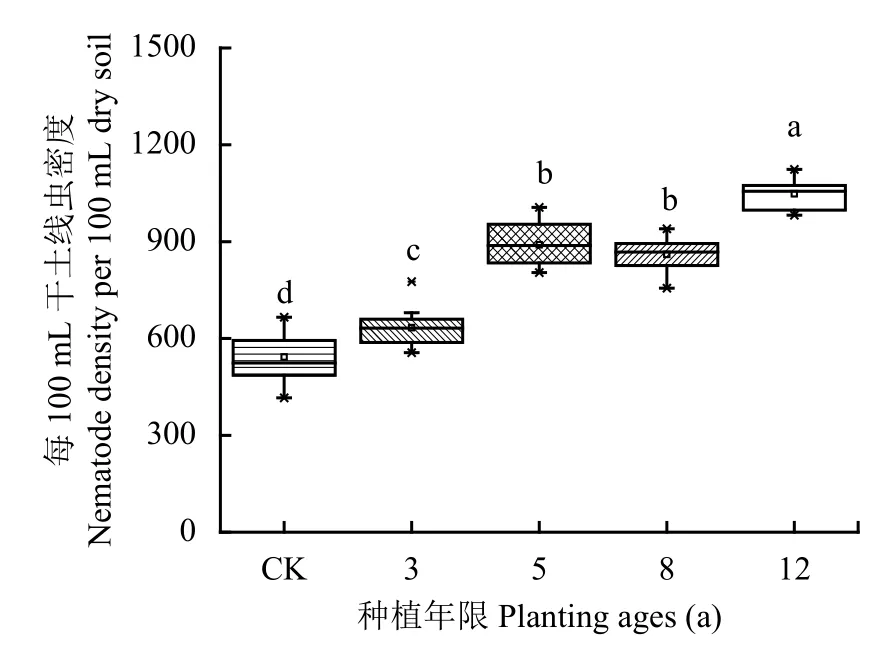
图1 不同种植年限条件下土壤线虫密度Fig. 1 The density of soil nematode in different planting ages
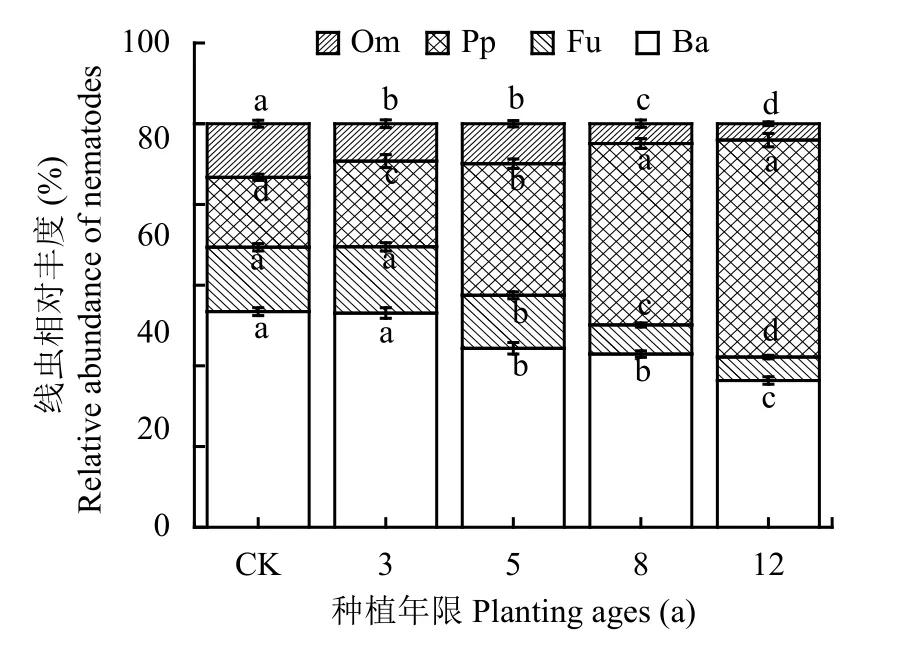
图2 不同种植年限条件下土壤线虫营养类群相对丰富度Fig. 2 The relative abundance of soil nematode trophic groups in different planting ages
土壤线虫营养类群相对丰度变化如图2所示。在所有样地中,食细菌线虫均为优势类,占个体总数的26.17%~59.25%。与CK相比,种植年限5年、8年、12年显著降低了食细菌线虫比率 (P <0.01),分别降低了17.03%、19.68%和31.92%。植物寄生类线虫,占个体总数的12.33%~59.17%;土壤植物寄生类线虫比率各种植年限表现为12年 >8年 > 5年 > 3年 > CK,与 CK 相比,种植蔬菜3年、5年、8年、12年显著增加了土壤植物寄生类线虫比率 (P < 0.01),分别增加了43.53%、209.82%、311.38%、501.71%。食真菌线虫,占个体总数的3.61%~20.89%;土壤食真菌线虫比率各种植年限呈现为 CK > 3 年 > 5 年 > 8 年 > 12 年,其中种植 5、8、12年后显著降低了食真菌线虫的比率 (P < 0.01),分别降低了17.67%、54.79%和63.74%。杂食/捕食性线虫所占比率较低,占个体总数的和2.34%~18.08%,杂食/捕食性线虫的比率各种植年限表现为CK > 3 年 > 5 年 > 8 年 > 12 年,种植 3、5、8、12年显著降低了捕食性线虫的比率 (P < 0.01),分别降低了30.12%、25.29%、62.99%和69.40%。由此可见,研究区设施菜地土壤植食性线虫和食细菌线虫为优势种群,随着设施菜地种植年限增加,土壤植食性线虫比率增加,食细菌、食真菌和杂食/捕食性线虫比例降低。
表2表明,不同种植年限土壤线虫群落组成也发生了变化。在60个样地共分离得到土壤线虫4371条,43个属,其中似杆线虫属 (Rhabditoides),头叶属(Cephalobus),短体属 (Pratylenchus) 均为优势属,而真滑刃属 (Aaphelenchus) 和中矛线属 (Mesodorylaimus)为CK、3年和5年样地的优势属,根结线虫属(Meloidogyne) 为5、8和12年样地的优势属,螺旋属 (Helicotylenchus) 为12年样地的优势属,随种植年限增加,异皮属 (Heterodera) 线虫所占比率也逐渐增加。在获得的所有线虫中,cp2个体和种类最大,其次是cp3个体和种类数量,而cp5 的个体数和种类量最少。随着种植年限的增加土壤根结线虫属、螺旋属、异皮属线虫逐渐增加,造成植物寄生性线虫的危害逐渐增加,其中土壤根结线虫病最为严重。
采用线虫多样性指数 (H′)、均匀度指数 (J ′ )、物种丰富度 (SR) 来评价线虫群落的多样性。H′ 和J′指数各种植年限表现出 5 年 > 8 年 > 3 年 > CK > 12 年,与CK相比,种植年限5年和8年显著增加了土壤线虫H′ 指数 (P < 0.05),而种植年限12年显著降低了土壤线虫H′ 指数 (P < 0.05),种植年限5年显著增加了土壤线虫J′ 指数 (P < 0.05),而SR指数各种植年限表现为 5 年 > 3 年 > 8 年 > CK > 12 年,与 CK 相比,种植年限12年显著降低了土壤线虫SR指数 (P < 0.05),而种植年限5年显著增加SR指数 (P < 0.05),这表明随着种植年限的增加土壤线虫多样性和群落结构稳定性先增加后降低,尤其是种植年限12年降低明显 (表 3)。
自由生活线虫成熟度MI指数、线虫通路比值NCR指数、植物寄生线虫成熟度PPI指数、瓦斯乐斯卡WI指数表示不同处理土壤线虫群落功能结构特征 (表3)。MI指数各种植年限表现出CK > 3年 >5年 > 8年 > 12年,与CK相比,种植年限5年、8年和12年显著降低了MI指数 (P < 0.05);线虫PPI指数表现出相反的趋势,各种植年限呈现出12 年 > 8 年 > 5 年 > 3 年 > CK,与 CK 相比,种植3年、5年、8年和12年显著增加量PPI指数 (P <0.05),表明样地土壤生态系统受种植年限的干扰较大。WI指数各种植年限表现出CK > 3年 > 5年 >8年 > 12年,与CK相比,种植年限3年、5年、8年和12年显著降低了WI指数 (P < 0.05),其中种植年限12年样地的WI值 < 1,表明种植年限12年后土壤矿化途径主要由植物主导,土壤处于不健康状态。所处理样地的NCR值都大于0.5,表明土壤有机质的分解途径均以细菌为主,随着种植年限增加土壤有机质的分解途径没有改变 (表3)。
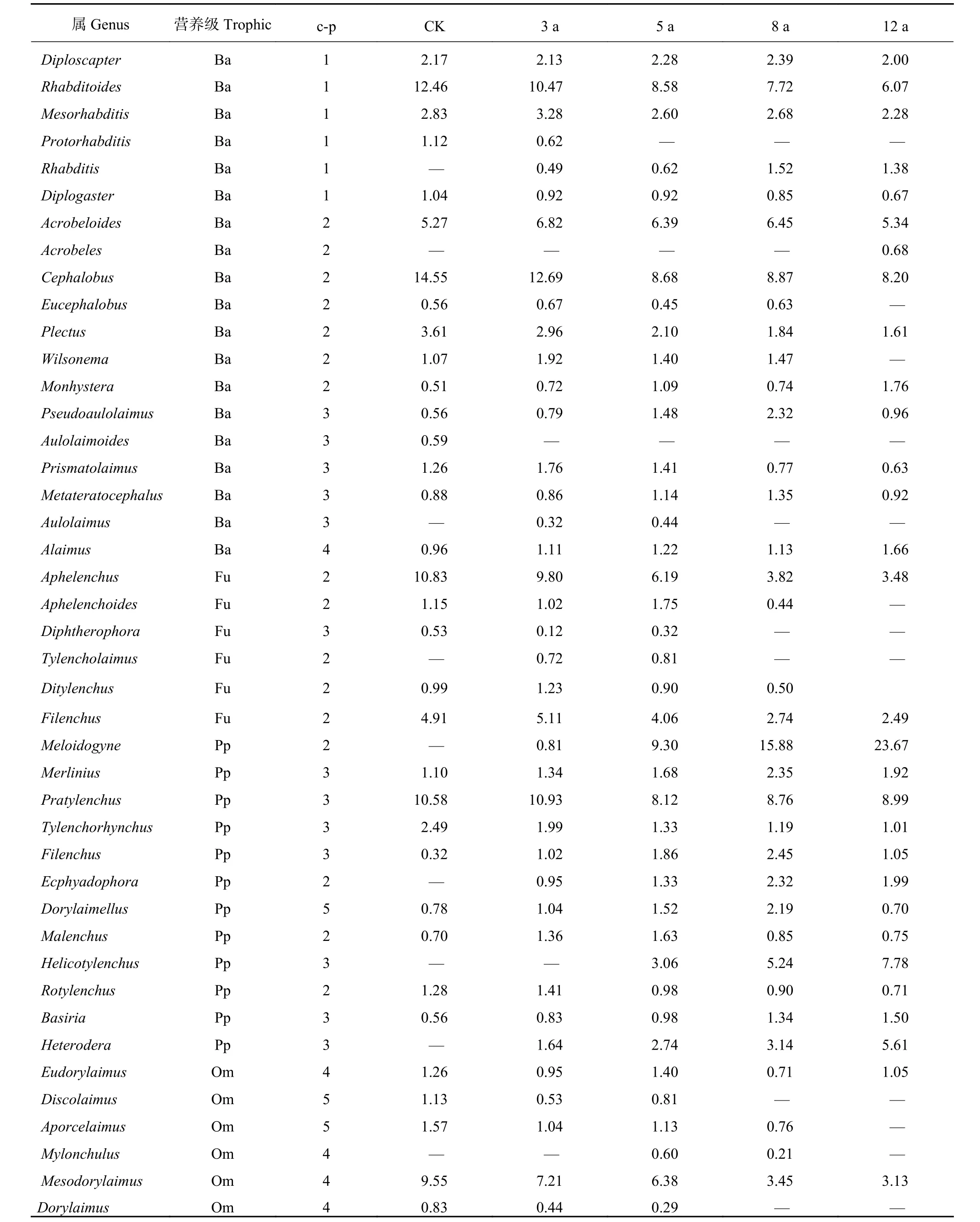
表2 不同种植年限土壤线虫群落组成及营养类群c-p值 (%)Table 2 Compositions of nematode community and c-p values of trophic groups in planting ages
2.3 土壤细菌和真菌的数量
图3显示,细菌和真菌数量各种植年限均表现为 5 年 > 12 年 > 8 年 > 3 年 > CK。土壤细菌的数量范围为 8.04 × 108~1.57 × 109,与 CK 相比,种植年限5年、8年、12年显著增加了土壤细菌的数量 (P < 0.01),分别增加了94.87%、33.70%和43.40%。土壤真菌的数量范围为 1.81 × 106~5.75 × 106,与 CK 相比,种植年限3年、5年、8年、12年显著增加了土壤真菌的数量 (P < 0.01),分别增加了27.73%、218.36%、70.53%、105.42%。随着设施菜地种植年限增加,土壤细菌和真菌的数量均表现出增加趋势,种植5年时达到最高。
2.4 土壤生物的影响因素分析
冗余分析表明,土壤理化性质变化是导致土壤线虫和微生物数量在不同种植年限存在差异的重要原因,8个环境变量解释了51%的数据总差异,其中第一轴和第二轴的解释率为41.4%和7.0%。土壤pH和容重 (BD) 与土壤线虫、细菌和真菌数量之间呈现负相关关系;土壤EC值、、有机碳 (SOC)、总孔隙度 (SK)、、与土壤线虫、土壤细菌和真菌数量之间呈现正相关关系 (图4)。根据箭头的长短可知,对土壤线虫数量影响程度最大的因素是土壤pH,最小的是。蒙特卡罗检验结果表明,pH (F=69.5,P < 0.001),BD (F=25.7,P < 0.001),EC (F=20.1,P < 0.001),(F=12.8,P < 0.01),SOC (F=7.9,P < 0.01),(F=7.2,P < 0.05),SK (F=6.3,P < 0.05),(F=5.7,P < 0.05),对土壤线虫数量影响达到显著水平。由此可见,土壤pH、孔隙度、容重、EC、、有机碳、是决定设施菜地土壤线虫、细菌和真菌数量变化的关键因子。不同种植年限改变土壤理化性质进而改变了土壤线虫与微生物数量。

表3 不同种植年限土壤线虫生态指数Table 3 Nematode ecological indices in different planting ages
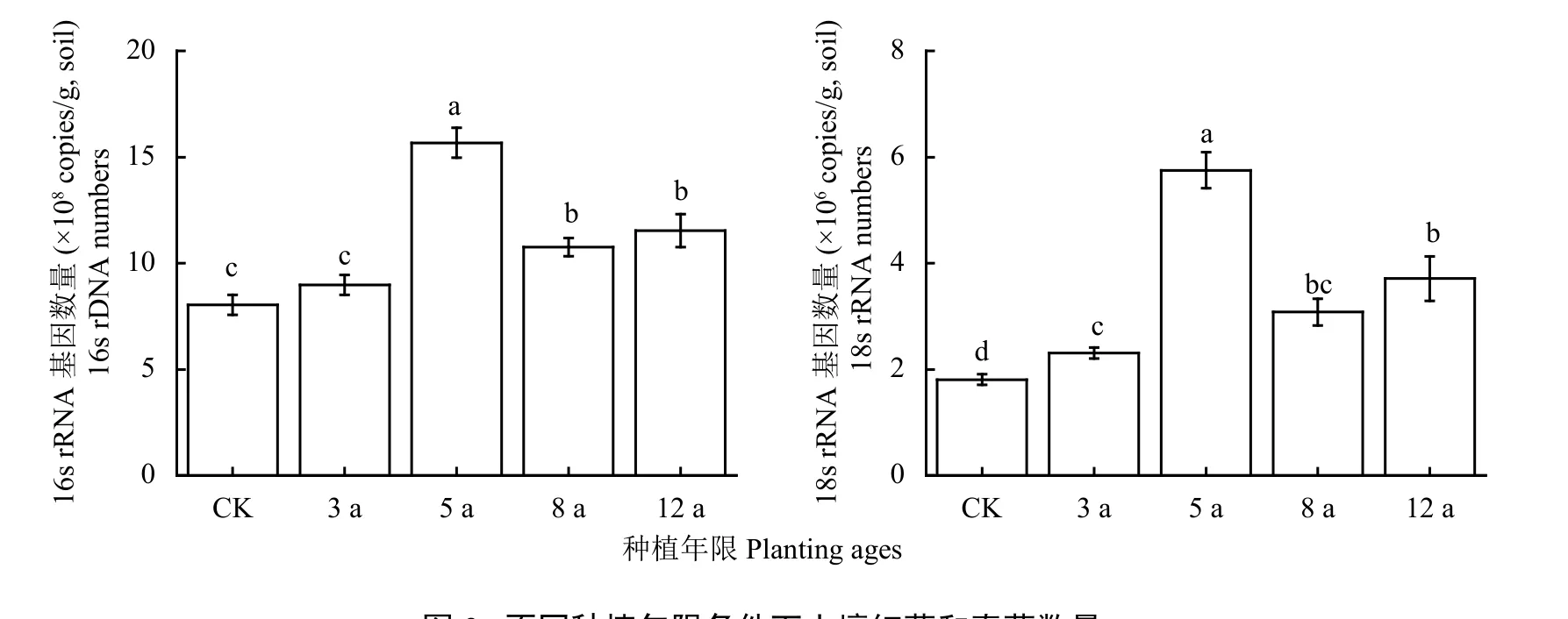
图3 不同种植年限条件下土壤细菌和真菌数量Fig. 3 The population of soil bacteria and fungi in different planting ages
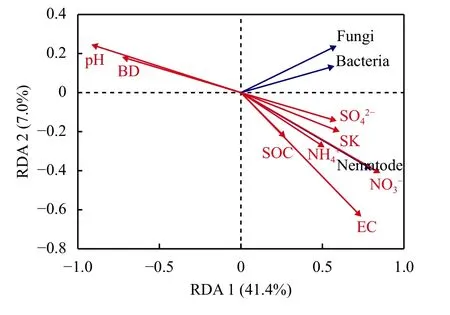
图4 土壤理化性质与土壤生物数量关系的RDA分析Fig. 4 The RDA ordination diagram of soil physical and chemical properties and number of soil organisms
2.5 微生物与食微线虫的关系
由土壤食微线虫与土壤微生物数量的关系 (图5)可知,所示。食细菌线虫与土壤细菌的数量呈现显著线性正相关关系 (R2=0.664,P < 0.001);而土壤食真菌线虫与真菌数量呈现二次函数关系,关系密切(R2=0.55,P < 0.01)。因此,设施菜地土壤食细菌线虫和食真菌线虫数量变化与土壤细菌和真菌的数量密切相关。
3 讨论
3.1 不同种植年限设施菜地土壤理化性质变化
土壤的理化性状是评价土壤生态环境质量的重要指标之一。随着种植年限的增加,北京设施菜地土壤有机质、、、、Cl–、、K+、Na+、Ca2+、Mg2+离子浓度均升高。这主要是因为:1) 设施蔬菜生产过程中化肥大量施用和较低的养分利用率;2) 设施蔬菜生产环境的封闭性、高温、高湿、少雨的条件下,导致淋洗程度低,使得大部分养分残留于土壤中并逐年累积,造成设施菜地土壤养分和离子含量较高[28–29]。随着种植年限的延长,北京设施菜地土壤酸化越明显,连续种植12 年的设施大棚,土壤pH值由7.10下降至6.18,下降了0.54个单位,这与高新昊等[28]、周德平等[30]的研究结果基本一致。pH值降低的主要原因可能是随着种植年限的延长,施用大量肥料导致、、Cl–等离子在土壤中残留促进土壤酸化[2,28],导致土壤环境恶化[31]。这说明在调查区的种植模式下,随着种植年限的延长,土壤出现了次生盐渍化。设施菜地随使用年限增加土壤容重有下降趋势,而土壤孔隙度有增加的趋势,这与曾希柏等[29]和高慧等[32]的研究结果基本一致,这与设施蔬菜生产过程中施加大量有机底肥密切相关。

图5 土壤微生物与食微线虫数量的关系Fig. 5 The relationship between the population of soil microbe and microbivorous nematode
3.2 不同种植年限设施菜地土壤微生物数量变化
土壤微生物不仅是土壤有机质和养分转化与循环的驱动力,也在土壤养分的转化与供应中起着重要作用,是评价土壤肥力水平的关键指标之一[33]。本研究利用荧光定量PCR分析了不同种植年限土壤细菌和真菌数量,发现设施菜地土壤中的细菌和真菌数量显著高于露天样地,这与宋蒙亚等[18]和王亚男等[19]的研究结论一致。这可能是由于:1) 设施菜地土壤中的有机质和养分含量高,可为微生物生长提供充足碳源和丰富底物;2) 设施菜地高温、高湿的土壤环境,特别适宜各类土壤微生物的生存和繁殖。设施菜地种植5年时土壤细菌和真菌数量均最高,然而种植8年和12年时土壤细菌和真菌数量又逐渐降低,这可能与土壤pH和土壤盐度变化有关,种植5年时pH表现为中性,而种植8年和12年时土壤pH变为酸性。通过对微生物数量与环境因子相关性分析 (RDA) 得出,土壤细菌和真菌数量与pH呈负相关关系,这与杨亚东等[34]研究结果一致。设施菜地施肥在一定时间内能够明显提高土壤微生物数量,改变微生物群落结构,然而若是继续大量施用化肥,导致速效养分含量显著增加,导致土壤较高的盐度和土壤严重酸化,高盐度和低pH降低了微生物的繁衍能力,因而,降低土壤微生物数量[34–35]。
3.3 不同种植年限设施菜地土壤线虫群落变化
线虫群落变化主要受资源有效性及所处生境微环境变化的调控[33,36],因而土壤线虫的群落动态可反映土壤的健康状况。本研究结果表明食细菌类线虫和植物寄生性线虫是研究区的主要营养类群,而杂食/捕食类线虫和食真菌类线虫的比例较低,土壤线虫cp2类群均为优势类群,生活策略以r对策为主,这与王殿东等[37]的研究结果一致。设施菜地不同种植年限下土壤线虫数量、群落组成和营养类群结构都存在一定的差异,并且不同营养类型的线虫对种植年限的响应不同。这些差异可能与不同线虫的生活史、食物资源和土壤健康状况的变化都有相关性[38–40]。
从线虫数量和类群上来看,随着种植年限延长土壤线虫密度增加,种植12年的设施菜地土壤由于根结线虫的大量增加而导致线虫数量显著高于其他种植年限,这与张雪艳等[12]的研究结果一致。这是由于随着种植年限增加,改变了线虫的食物来源及生存环境[37]。本研究线虫数量与环境因子的相关分析也证实这一观点,土壤线虫数量与土壤孔隙度、土壤养分、细菌数量之间呈现显著正相关关系而与土壤pH、土壤容重呈现负相关关系。这是由于:1) 土壤中有机质、养分以及植物根系在土壤中积累,为土壤线虫提供了更多的食物来源和庇护所;2) 土壤理化性质的改变在一定程度上增加了土壤细菌数量,为食细菌线虫提供了直接的食物来源;3) 由于土壤线虫本身没有自主运动能力,其活动依赖于土壤孔隙水进行;随着种植年限延长土壤孔隙度得到改善,且设施菜地高湿的环境使得土壤孔隙度含水量提高,利于土壤线虫的运动及捕食,扩大其群落规模;4)蔬菜连作种植模式以及高温和高湿的环境使得植物寄生线虫尤其是根结线虫大爆发,进而增加线虫总数[37–38]。对其他线虫类群而言,随着种植年限延长土壤食物网中食细菌、真菌线虫和杂食/捕食性的线虫比率随之降低,种植12年时线虫营养类群结构失调,这可能是由于这些类群线虫与植物寄生性线虫竞争力逐渐降低[39–40]。由于杂食/捕食性线虫是决定土壤食物网复杂性的关键生物类群,因此随着种植年限增加土壤食物网复杂性降低,可能影响到土壤微生态平衡[41]。
土壤线虫群落的多样性和均匀性变化也呈现为先增加后降低的趋势,这与张雪艳等[12]的研究结果类似。尤其是种植12年时设施菜地物种数、多样性和均匀性都降低,表明种植12年群落趋于单一化,多样性和稳定性较差,说明种植12年时设施菜地土壤生态系统发生退化;而种植5年时设施菜地物种数、多样性和均匀性都最高,这可能由于种植5年土壤理化性质和细菌、真菌数量为土壤线虫群落组成提供稳定的生态环境因素[40]。种植5年是土壤生态系统发生变化的转折点。
自由生活线虫成熟度指数WI,常被用于分析土壤食物网的矿化途径和评价土壤健康状况[4,42,27],种植12年的设施菜地土壤线虫的WI < 1,这表明此种植年限土壤矿化途径从植物到植食性线虫,土壤处于不健康状态;种植3年、5年和8年的MI > 1,这表明此时土壤矿化途径是由食细菌性线虫和食真菌性线虫参与,土壤处于相对健康状态。设施菜地连续种植使矿化途径的主要参与者由食微线虫转变为从植物到植食性线虫为主。线虫通路比值NCR指数,常用于表征腐屑食物网有机质的分解途径[42,27]。本研究中NCR处于 0.76~0.86 之间,均高于 0.5,说明不同种植年限的土壤食物网以食细菌通道为主,土壤有机质以细菌分解途径为主,这与张雪艳等[12]、王殿东等[37]的研究结果一致。自由生活线虫的成熟度MI指数和植物寄生线虫的成熟度PPI指数表征土壤线虫群落功能结构特征,亦可用以评价土壤线虫群落受干扰的程度[27]。在本研究中,MI指数随种植年限增加而逐渐降低,在种植12年时达到最低,而PPI指数表现出相反的趋势,这表明随种植年限增加土壤生态系统受干扰程度也随之增加,种植12年土壤生态系统扰动最强烈,降低了土壤生态系统的稳定性。
4 结论
1) 设施菜地土壤理化性质变化与种植年限有密切关系。随着种植年限的增加,设施菜地土壤有机碳、Ca2+、Mg2+离子浓度均升高,土壤pH逐渐降低,土壤孔隙度逐渐增加。
4) 种植12年的菜地土壤线虫总数显著高于其他年限,但群落趋于单一化,多样性和稳定性较差。
5) 研究区域的设施菜地土壤食物网以食细菌通道为主,土壤有机质以细菌分解途径为主。随着种植年限延长,WI、MI指数逐渐降低,而PPI指数逐渐增加,其中种植12年设施土壤矿化途径由食微线虫参与为主转变从植物到植食性线虫为主要参与者,随种植年限增加土壤生态系统受干扰程度也随之增加,连续种植12年的菜地土壤生态系统扰动最强烈,土壤健康状况变差。
