转录组测序技术筛选安徽乌菜与普通白菜的差异表达基因
2018-12-06王明霞田红梅江海坤
王明霞 田红梅 江海坤 贾 利 王 艳 方 凌 *
(1安徽省农业科学院园艺研究所,安徽合肥 230031;2园艺作物种质创新与生理生态安徽省重点实验室,安徽合肥 230031)
乌 塌 菜〔Brassica campestrisL. ssp.chinensis(L.)Makinovar.rosularisTsen et Lee〕别名塌菜、塌棵菜、黑菜等,2n=20,是十字花科芸薹属芸薹种白菜亚种的一个变种,起源于南北气候区之间。安徽乌菜是乌塌菜的一种,其植株形态介于普通白菜与大白菜之间,是安徽省的著名特色蔬菜之一,具有很多优良性状和独特的商品性,如抗寒性、抗病性、耐抽薹、叶面皱泡、口感与风味等,对作物遗传改良及研究白菜类的进化与分化具有重要价值,是宝贵的种质资源(李曙轩,1990)。曹家树和曹寿椿(1995)认为环境条件决定着植物演化的方向,中国白菜各类群的变异主要是叶部性状的变化,从叶片等性状入手探讨其演化关系是有意义的。叶面上有皱泡是安徽乌菜区别于其他白菜亚种的一个典型特征,前期生产实践表明,皱泡多而密的安徽乌菜耐寒性较强,商品性好。但是,目前对安徽乌菜形态变异的分子机制知之甚少,这在很大程度上妨碍了对安徽乌菜叶部形态发生过程的深入了解,使许多生产上的现象无法得到正确的解释和证实。为了研究安徽乌菜与普通白菜叶部形态上的差异,本试验拟在转录组水平上研究安徽乌菜与普通白菜的差异表达基因,揭示其转录组序列水平和转录组表达量水平的变异,发掘安徽乌菜叶部形态变异的转录组差异和分子机制,为安徽乌菜基因挖掘和进一步开发优异性状材料提供理论基础和技术支撑,同时也为研究白菜类蔬菜进化及性状建成机理提供理论依据。
1 材料与方法
1.1 试验材料
以安徽省农业科学院园艺研究所自育的普通白菜自交系C-14-007、安徽乌菜自交系黄心乌1408和黑心乌1422为试材,2016年9月在安徽省农业科学院园艺研究所试验地进行播种,播后30 d定植,定植前每667 m2施腐熟有机肥3 000~4 000 kg、三元复合肥(N-P-K为17-8-20)25~30 kg,深翻耙平后做畦。当第15片叶展开时,选取生长发育一致的植株,取第15片叶迅速放入液氮中,每份材料取2份样品作为重复,保存至-80 ℃冰箱备用。普通白菜C-14-007的2份样品编号为T01、T02,黄心乌1408的2份样品编号为T03、T04、黑心乌1422的2份样品编号为T05、T06。基于Illumina HiSeq2500测序技术平台,利用双末端测序(Paired-End)的方法,构建转录组文库进行双末端测序,获得转录组测序数据。
1.2 试验方法
1.2.1 RNA提取与检测 采用天根生化科技(北京)有限公司的RNA提取试剂盒提取安徽乌菜自交系1408、1422和普通白菜C-14-007第15片叶的总RNA,分别采用Nanodrop分光光度计、Qubit 2.0荧光计、Aglient 2100生物分析仪检测RNA样品的纯度、浓度和完整性等,以保证使用合格的样品进行转录组测序。
1.2.2 RNA文库构建、RNA-Seq及文库质控
RNA样品检测合格后,利用Illumina’s TruSeq RNA Sample Preparation Kit(Illumina Inc,San Diego,CA,USA)构建mRNA-Seq文库。mRNA的纯化、片段打断、cDNA合成、加接头、PCR扩增以及RNA-Seq均在北京百迈客生物科技有限公司完成。文库构建完成后,分别使用Qubit 2.0荧光计和Agilent 2100生物分析仪对文库的浓度和插入片段大小(insert size)进行检测,使用Q-PCR方法对文库的有效浓度进行准确定量,以保证文库质量。
1.2.3 数据组装 去掉低质量的reads,利用Trinity软 件(http://trinityrnaseq.sourceforge.net/) 对 clean reads进行组装。首先将测序reads打断为较短的片段(K-mer),然后将这些小片段延伸成较长的片段(contig),并利用这些片段之间的重叠,得到片段集合(component),最后利用de Bruijn图的方法和测序reads信息,在各个片段集合中分别识别转录本序列。
1.2.4 差异表达基因的筛选 FPKM(fragments per kilobase of transcript per million mapped reads)是每百万reads中来自比对到某一基因每千碱基长度的reads数目,是转录组测序数据分析中常用的基因表达水平估算方法。FPKM能消除基因长度和测序量差异对计算基因表达的影响,计算得到的基因表达量可直接用于比较不同样品间的基因表达差异。在筛选过程中,将FDR(false discovery rate)<0.01且差异倍数FC(fold change)≥2作为筛选标准,其中FC表示两样品(组)间表达量的比值。普通白菜与黄心乌差异基因以G1表示,普通白菜与黑心乌的差异基因以G0表示。
1.2.5 差异表达基因功能注释 使用BLAST软件将差异表达基因序列与nr(http://www.ncbi.nlm.nih.gov/)、GO(http://www.geneontology.org/)、COG(http://www.ncbi.nlm.nih.gov/COG/)、KEGG(http://www.genome.jp/kegg)数据库比对,获得注释信息。
2 结果与分析
2.1 转录组测序及数据组装
利用转录组测序技术对普通白菜和安徽乌菜(黄心乌、黑心乌)进行分析,通过转录组测序共获得27.75 Gb clean data,各样品测序数据量均达到4.00 Gb,Q30碱基百分比在90.53%以上。由表1可知,通过组装共得到127 988条转录本和46 950条unigene,平均长度分别为1 327.71 bp和856.26 bp,转录本与unigene的N50长度分别为1 857 bp和1 505 bp,而≥1 000 bp的unigene共13 076条(27.85%)。
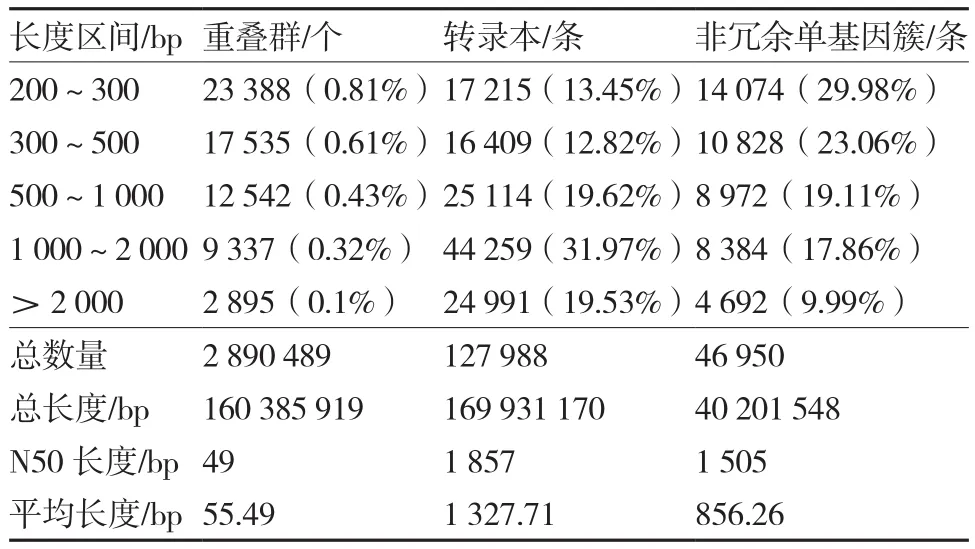
表1 转录组测序数据组装结果
2.2 差异基因筛选
以FDR<0.01且FC≥2作为筛选标准,筛选到普通白菜与黄心乌的差异基因(G1)共3 191个,普通白菜与黑心乌的差异基因(G0)共2 180个,G1与G0共同的差异基因有1 296个。
2.3 差异表达基因功能注释
将筛选到的1 296个共同差异表达基因与COG、GO、KEGG、KOG、Pfam、Swiss-Prot、nr等数据库进行比对,共有1 156个差异表达基因被注释,其中223个差异表达基因注释到COG数据库,864个差异表达基因注释到GO数据库,200个差异表达基因注释到KEGG数据库,445个差异表达基因注释到KOG数据库,716个差异表达基因注释到Pfam数据库,785个差异表达基因注释到Swiss-Prot数据库,1 150个差异表达基因注释到nr数据库。
注释到COG数据库中的差异表达基因有223个,占差异表达基因总数的17.21%,这些基因按功能进行分类,共分为21类,其中一般功能预测类基因共72个(32.29%),其次是与转录相关的差异基因有36个(16.14%),复制、重组及修复类功能基因共35个(15.70%),而核苷酸转运和代谢功能相关基因有3个(1.35%)(图1)。
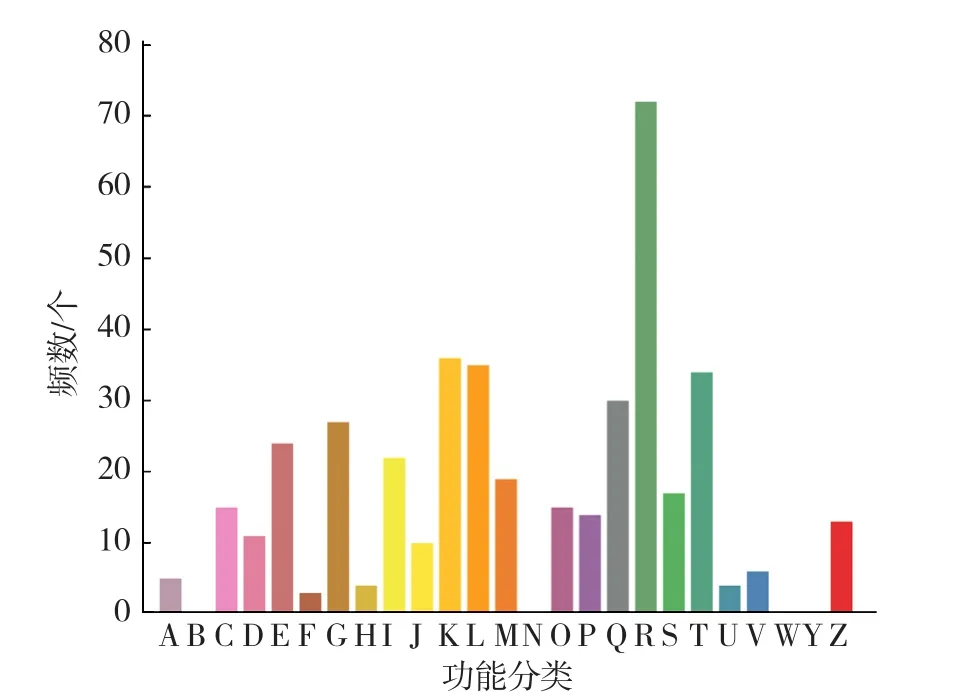
图1 安徽乌菜与普通白菜差异表达基因COG数据库功能注释
注释到GO数据库中的差异表达基因有864个(66.67%),分为细胞组成、生物过程、分子功能3大类53个分支,其中细胞组成类基因共764个,生物过程类基因共707个,分子功能类基因共595个,细胞组成、生物过程、分子功能全包含的差异表达基因486个。在细胞组成类基因中,与细胞组分相关的基因和与细胞相关的基因均为597个;在分子功能类基因中,与结合相关的基因380个,其次是与催化活性相关的基因357个;在生物过程类基因中,与细胞过程相关的基因、与代谢过程相关的基因及与单个有机体过程相关的基因均为441个(图 2)。
KEGG数据库能够系统分析基因产物在细胞中的代谢途径以及功能。将普通白菜与黄心乌的差异表达基因和普通白菜与黑心乌的差异表达基因与KEGG数据库进行比对,结果有200个差异表达基因被注释到80条通路,这些通路分别参与遗传信息过程、有机系统、环境信息过程、代谢和细胞过程。在这些差异表达基因中,参与植物病原菌互作通路的最多(ko04626,22个),其次是参与植物激素信号转导通路(ko04075,19个)和类苯基丙烷生物合成通路(ko00940,11个)(图3);而参与油菜素类固醇生物合成(ko00905)、单萜类生物合成(ko00902)等通路的差异表达基因只有1个。在植物激素信号转导通路(ko04075)中,与生长素合成相关的基因有4个,与玉米素生物合成相关的基因有2个,与油菜素内酯合成相关的基因有2个。

图2 安徽乌菜与普通白菜差异表达基因GO数据库功能注释
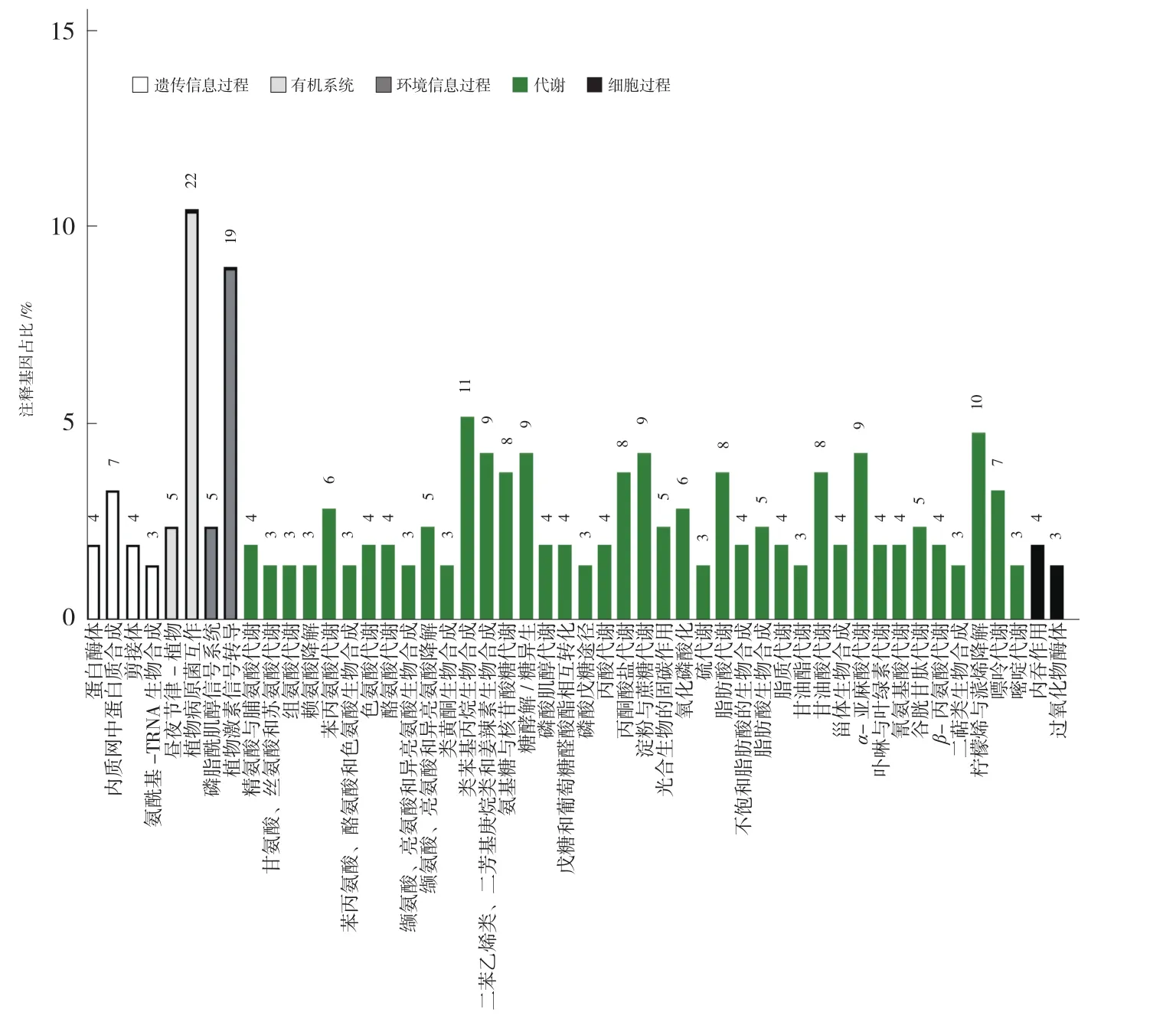
图3 安徽乌菜与普通白菜差异表达基因KEGG数据库功能注释
3 结论与讨论
叶片是植物光合作用的主要器官,叶片性状能够反映植物适应环境变化所形成的生存对策,是指示植物生理生态特征及对环境变化适应对策的有效指标(宋璐璐 等,2011)。叶面上有皱泡是安徽乌菜区别于其他白菜亚种的一个典型特征,叶面皱泡的产生增加了叶面积,直接影响了比叶面积(叶面积与叶片干质量的比值,SLA)大小,比叶面积是植物获取资源的衡量指标,在很大程度上可以解释植物光合作物、呼吸作用、叶寿命和潜在生长速率的种间变异(Ackerly et al.,2002)。笔者认为安徽乌菜是普通白菜从南到北过渡过程中为了适应环境变化而逐渐进化而成,因此安徽乌菜与普通白菜相比具有更强的抗寒性(Tian et al.,2018)。差异表达基因存在于植物不同生长发育期、不同组织、不同处理和不同品种间,可通过研究差异表达基因探讨生理变化的机制(Logacheva et al.,2011;Wang et al.,2013;Wu et al.,2014;Zhang et al.,2016)。本试验对安徽乌菜和普通白菜进行了转录组测序,获得安徽乌菜和普通白菜差异表达基因,为发掘安徽乌菜叶部性状变异的分子机制和研究白菜类蔬菜进化提供了一定的数据支持。
叶片发育受植物体内严格的遗传机制和体外环境因子的调控,植物激素尤其是生长素在协调体内、外调节机制中起着不可或缺的作用,越来越多的研究表明生长素的稳态调控、剂型运输和信号转导与叶片发育和形态建成具有紧密联系,暗示生长素在叶片生长发育过程中具有关键的调控功能(李林川和瞿礼嘉,2006)。Wang等(2011)研究发现,白菜在基因组发生加倍之后,与器官形态变异有关的生长素相关基因发生了显著的扩增,白菜基因组复制导致了许多与形态变异有关的基因存在更多拷贝,这可能是白菜类蔬菜具有丰富的根、茎、叶形态变异的根本原因。本试验通过对黑心乌、黄心乌与普通白菜进行差异表达基因筛选,共获得1 296个差异表达基因,200个差异表达基因注释到KEGG数据库,19个基因参与了植物激素信号转导通路,其中与生长素合成相关的基因有4个。1种激素往往参与调节发育过程的多个方面,多种激素也可以同时参与调控某一特定发育过程(Gray,2004)。如玉米素主要分布于进行细胞分裂的部位,对细胞的伸长有阻碍作用,控制顶端优势,促进细胞的横向扩张(杜旭升和蒲晓亚,2014);油菜素内酯调控细胞的伸长和细胞分裂,同时还可促进光合作用和提高抗逆性。本试验从参与植物激素信号转导通路的19个基因中发现了2个与玉米素合成相关的基因,2个与油菜素内酯合成相关的基因。这些结论将为下一步的基因发掘提供支持。今后将继续研究与植物激素、次生代谢相关的差异表达基因,为探讨安徽乌菜叶部形态变异分子机制提供一定的数据支持。
