青海春小麦品种‘青春38’成株期抗条锈性遗传解析
2018-12-05张调喜闫佳会侯璐张贵姚强马麟郭青云
张调喜 闫佳会 侯璐 张贵 姚强 马麟 郭青云


摘要
为明确青海春小麦品种‘青春38成株期抗条锈性的遗传基础,以‘青春38为父本与感病春小麦品种‘Taichung 29(T29)杂交构建F2∶3代分离群体。在青海西宁和互助两地田间病圃进行了抗条锈性鉴定,应用植物数量性状主基因+多基因混合遗传模型单个分离世代分析方法,解析‘青春38的抗条锈性遗传特点。结果表明,‘青春38/‘T29 F2∶3群体单株的病害严重度和反应型在两个试验点均未呈现连续性分布,也不符合正态分布,初步推测‘青春38对小麦条锈病的成株期抗性具有质量性状特征;以严重度或反应型数据进行遗传分析,‘青春38在两个试验点对小麦条锈病的成株期抗性表现的最优遗传模型均属2 对主基因遗传,只是主基因的作用方式(C-1:2MG-ADI加性-显性-上位性,C-4:2MG-EA 等加性,C-6:2MG-EEAD 等显性)有所不同。
关键词
春小麦; 青春38; 抗条锈性; 遗传模型
中图分类号:
S 435.121
文獻标识码: A
DOI: 10.16688/j.zwbh.2017416
Genetic analysis of adult plant resistance to stripe rust in Qinghai
spring wheat cultivar ‘Qingchun 38
ZHANG Tiaoxi YAN Jiahui HOU Lu ZHANG Gui YAO Qiang MA Lin GUO Qingyun1
(1. Academy of Agricultural and Forestry Sciences, Qinghai University, Qinghai Academy of Agricultural and Forestry Sciences,
Scientific Observing and Experimental Station of Crop Pests in Xining, Ministry of Agriculture, Key Laboratory of Agricultural
Integrated Pest Management,State Key Laboratory of Plateau Ecology and Agriculture, Xining 810016, China;
2. Datong Agricultural Technology Promotion Center of Xining, Qinghai Province, Xining 810100, China)
Abstract
In order to elucidate the genetic basis of adult plant resistance of spring wheat ‘Qingchun 38 to stripe rust in Qinghai, a F2∶3 segregating population was developed by crossing ‘Qingchun 38 (as male parent) with ‘Taichung 29 (T29) (as female parent), a susceptible cultivar. Disease screening was conducted at Xining and Huzhu in Qinghai Province. Single generation of major gene plus polygene mixed inheritance model was used to analyze the inheritance of stripe rust resistance in ‘Qingchun 38. Disease severity and infection type of ‘Qingchun 38/‘T29 F2∶3 populations showed discrete distributions, suggesting a qualitative character of stripe rust resistance in ‘Qingchun 38. Genetic analysis results in two environments indicated that the two major gene model could explain the resistance of ‘Qingchun 38 on disease severity and infection type, but the gene interactions (C-1: 2MG-ADI additive-major-epistatic interaction, C-4: 2MG-EA equal additive effect,C-6: 2MG-EEAD equal dominance effect) were different.
Key words
spring wheat; Qingchun 38; stripe rust resistance; inheritance model
小麦条锈病是由小麦条锈菌 Puccinia striiformis f.sp. tritici 引起的世界性小麦病害,是影响小麦生产最重要的病害之一,该病害在低温和潮湿环境下易发生,从小麦一叶期到成熟期只要植株仍然是绿色都可以发生侵染[1]。研究与实践证明,选育和利用抗病品种是防治小麦条锈病最经济、安全和有效的措施[1-3],然而,小麦条锈菌群体毒性结构复杂,新毒性小种不断形成,容易使小麦品种抗病性丧失,致使小麦条锈病周期性流行。不断发掘新的抗条锈病基因,实现抗病基因的多样化及合理布局是一项有战略意义的基础研究工作,抗条锈病基因发掘及其遗传特点研究是抗病育种的重要基础。
青海省是我国小麦条锈菌主要越夏流行区之一[4],青海省小麦种植垂直分布,春小麦和冬小麦交错种植,每年拥有大量的小麦条锈病越夏菌源,主要以晚熟春麦上的菌源为主,可以保持大量的菌源到早播冬麦出苗后,为我国东部冬麦区秋苗发病提供较多的有效菌源。同时青海省种植的春小麦中抗条锈病品种较少,主要以感病品种‘阿勃为主,给我国小麦条锈病越夏菌源区的区域治理造成非常不利的局面。
为增加抗病基因的多样性,国内研究者做了大量的工作,中国农业科学院植物保护研究所小麦病害团队对国内大量小麦农家品种和生产品种进行了系统的抗条锈性鉴定和研究[5-10]。曹世勤等[11]分别对甘肃省 26 个春小麦品种(系)进行了苗期抗条锈基因分析及成株期抗病性评价,此外还于2003-2013年对4 291 份冬小麦和710 份春小麦进行了系统的抗条锈性鉴定[12]。李永平等[13]对86份贵协系小麦种质资源进行了抗条锈性分析;王万军等[14]对临麦系列春小麦品种进行了抗条锈性评价;周新力等[15]将80份国外春小麦种质资源做了抗条锈病性评价;姚强等[16-17]也对120个春小麦品种(系)做了成株期抗条锈性评价,并对青海春小麦品种‘青春39进行了苗期抗条锈性基因遗传分析,侯璐等[18-19]对搜集的81份春小麦种质资源进行了系统的抗条锈性鉴定,并对4份春麦种质资源进行了苗期抗病基因遗传分析。国内就春小麦品种成株期抗条锈病基因的遗传分析研究相对较少。本研究组从2010-2016年连续7年对大田成株期抗条锈性进行观察,发现春小麦品种‘青春38一直表现高抗,是很好的抗源材料[17],本研究拟解析‘青春38在成株期的抗条锈病基因及遗传特点和抗性特点,促进其有效合理利用。
1 材料与方法
1.1 试验材料
‘青春38是青海春小麦主栽品种,由本研究室保存;感病对照春小麦品种‘Taichung 29 (T29)由中国农业科学院植物保护研究所徐世昌研究员提供,以‘青春38为父本与‘T29杂交、自交获得的F2∶3代群体,由本研究室保存。
1.2 方法
抗性鉴定于2015 年和2016年分别在青海省互助县威远镇锈病常发地青海省农林科学院植保所小麦条锈病抗性鉴定自然发病圃和青海省西宁市青海省农林科学院植保所小麦条锈病抗性鉴定自然发病圃进行。
每个亲本各播种1行,行长1 m,行距0.3 m;2015年F2群体单粒点播,每行播 10粒,每20 行加设1行 ‘Taichung 29作为感病对照,在西宁市种植的F2群体共有118个单株收获得到对应的118个F3家系种子,在互助县种植的F2群体共有156个单株收获得到对应的156个F3家系种子。2016年F3家系群体继续于同一地方种植测试,每个F3家系种1行,每20 行加设1行‘Taichung 29作为感病对照;鉴定圃四周各种3 行‘Taichung 29作诱发行。 播种时间为每年4月上旬,从 7月上旬开始(即当‘Taichung 29充分发病的时候)对亲本及F2群体(2015年)和每个F3家系(2016年)每个单株进行调查,记载反应型和严重度,调查2次,间隔7 d,反应型(IT)按 Line 等[3] 0~9 級的方法记录,田间病害严重度(DS,%)按0、2、5、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、 85、90、 95、100分级记录[15],选用2次调查中最大严重度作最终统计。
采用 Excel 2016 软件统计数据并绘制‘青春38/‘T29F2∶3群体在两个环境中的严重度和反应型的频率分布条形图;利用Minitab 16 软件做正态性检验分析;利用刘兵[20]研制的植物数量性状主基因+多基因混合遗传的分离分析软件(软件包SEA-F2)进行分析,用极大似然和期望最大化算法估计分布参数,AIC(Akaikes information criterion,赤池信息准则)值较小且适合性检验较优的模型为适宜遗传模型。采用最小二乘法从一阶分布参数估计一阶遗传参数,并估计主基因和多基因遗传方差和遗传率等二价遗传参数。
2 结果与分析
2.1 亲本及F2∶3群体条锈病抗性统计分析
2015和2016年在互助和西宁两个试验地点条锈病均发病比较充分。感病亲本‘Taichung 29的病害反应型(IT)均为9级,严重度(DS)为95%;抗病亲本‘青春38的反应型(IT)为3级,严重度(DS)为15%;F2群体2015年和对应的F3家系群体2016年分别在两个环境中反应型和严重度的频率分布如图1和图2。可知‘青春38/‘T29 F2∶3群体的IT和DS在2个环境下均未呈现连续性分布。
图1 ‘青春38/‘T29 F2∶3群体在互助试验地所测试反应型和病害严重度的频率分布条形图
Fig.1 Frequency distribution of infection type (IT) and disease severity(DS)of F2∶3 populations from
‘Qingchun 38/‘T29 tested in Huzhu
图2 ‘青春38/‘T29 F2∶3群体在西宁试验地所测试反应型和病害严重度的频率分布条形图
Fig.2 Frequency distribution of infection type (IT) and disease severity(DS)of F2∶3 populations from
‘Qingchun 38/‘T29 tested in Xining
‘青春38/‘T29 F2群体在两个环境中反应型和病害严重度的正态概率图如图3。由正态概率图可以看出,两个杂交组合F2群体在2个环境下DS和IT值的正态性分布检验结果均为P<0.005,说明‘青春38/‘T29 F2群体在两个环境中DS和IT值均不符合正态性分布。
综合抗性统计分析结果初步推测,‘青春38对小麦条锈病成株期抗性均具有质量性状抗性特征。
2.2 遗传模型的选择
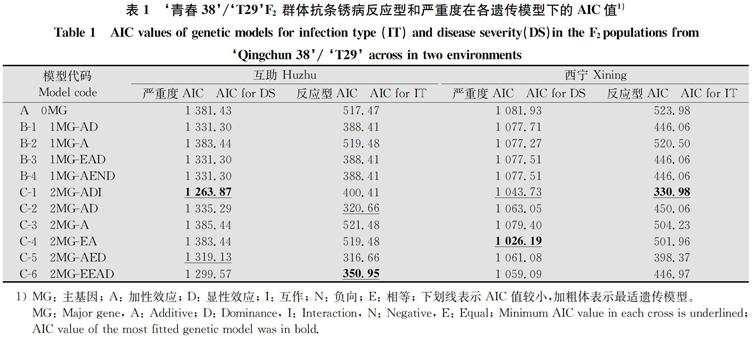
用主基因+多基因混合遗传的F2群体分析方法对‘青春38的成株期抗条锈性基因进行分析(表 1),得到0 对主基因(A)、1 对主基因(B)和2 对主基因(C) 等 3类11 种遗传模型的AIC值。选取最小AIC值与最小AIC 值比较接近的遗传模型作为候选模型,对备选模型进行适合性检验,包括均匀性检验、 Smirnov 检验和 Kolmogorov检验,获得U12、U22和U32、nW2和Dn统计量,选择达到显著水平的检验个数最少的模型为最适遗传模型。
2.2.1 ‘青春38的成株期抗条锈性基因遗传模型的选择
根据在互助测试的分析结果,用严重度数据分析时,C-1、C-6和C-5的AIC值最低,分别为1 263.87、1 299.57和1 319.13;适合性检验结果(省略)显示,这3个模型统计量未达到显著水平个数均为1个,而C-1模型的AIC值最小,因此,C-1模型可以作为最适遗传模型。用反应型数据分析时,C-5、C-2和C-6的AIC值最低,分别为316.66、320.66和350.95;结合适合性检验结果(数据省略),达到显著水平的检验个数最少的模型C-6,可以作为最适遗传模型。
图3 ‘青春38/‘T29 F2群体在互助(a)和西宁(b)试验地反应型和病害严重度的正态概率图
Fig.3 Normal probability plot of infection type (IT) and disease severity(DS)of F2 populations from
‘Qingchun 38/‘T29 in Huzhu (a) and Xining (b)
在西宁测试的分析结果,用严重度数据分析时,C-4、C-1和C-6的AIC最低,分别为1 026.19、1 043.73和1 059.09;适合性检验结果(省略)显示,这3个模型统计量均未达到显著水平,而C-4模型的AIC值最小,作为最适遗传模型。用反应型数据分析时,C-1和C-5的AIC值最低,且值相对比较接近,分别为330.98和398.37;适合性检验结果(省略)显示,这2个模型统计量未达到显著水平个数均为1个,而C-1模型的AIC值最小,因此,C-1模型作为最适遗传模型。
由此可见,在两个地点,选用严重度数据或反应型数据分析最优遗传模型均为2对主基因遗传,只是主基因的作用方式(C-1:2MG-ADI为加性-显性-上位性,C-4:2MG-EA为等加性,C-6:2MG-EEAD为等显性)有所不同。
表1 ‘青春38/‘T29F2群体抗条锈病反应型和严重度在各遗传模型下的AIC值1)
Table 1 AIC values of genetic models for infection type (IT) and disease severity(DS)in the F2populations from
‘Qingchun 38/ ‘T29 across in two environments
1) MG: 主基因; A: 加性效應; D: 显性效应; I: 互作; N: 负向; E: 相等; 下划线表示AIC 值较小,加粗体表示最适遗传模型。
MG: Major gene, A: Additive; D: Dominance, I: Interaction, N: Negative, E: Equal; Minimum AIC value in each cross is underlined; AIC value of the most fitted genetic model was in bold.
2.2.2 最适遗传模型相关参数估计
根据最适遗传模型估计调查发病反应型和严重度的一阶和二阶遗传参数(表2)。‘青春38/‘T29F2群体在互助测试,‘青春38成株期抗条锈性以严重度数据分析为C-1模型,即2对加性-显性-上位性主效基因作用,2对主基因(A和B)加性效应均表现为正向效应(da>0,db>0),显性效应表现为1个负向效应,1个正向效应(ha<0,hb>0),2对主基因间加性×加性互作为负向(i<0),其他互作均为正向作用;以反应型数据分析为C-6模型,即2对等显性主基因作用,加性效应表现为1个正向效应(da>0)。在西宁测试,以严重度数据分析为C-4模型,即2对等加性主基因作用,表现为1个正向加性效应(da>0);以反应型数据分析结果为C-1模型,即2对加性-显性-上位性主基因作用,2对主基因(A和B)加性效应和显性效应均表现为正向效应(da>0,db>0;ha>0,hb>0),基因间加性×加性互作为负向(i<0),其他互作均为正向。在互助试验地条件下,主基因遗传率为97.92%和86.11%,西宁试验地条件下,主基因遗传率为95.10%和99.15%。说明在不同环境下,‘青春38成株期抗条锈性均具有很高的遗传力,而且主基因的遗传率差异不大,环境因素对主基因的影响较小,遗传稳定性较高。
表2 各F2群体在最优遗传模型下遗传参数的估计值1)
Table 2 Genetic parameters estimated from the best models in the F2 populations
1) m: 群体平均值; da: 第1对主基因加性效应; db: 第2 对主基因加性效应; ha: 第1对主基因显性效应; hb: 第2对主基因显性效应; i: 2对主基因间加性×加性互作; jab: 2对主基因间加性×显性互作; jba: 2对主基因间显性×加性互作; l: 2对主基因间显性×显性互作; σ2mg: 主基因遗传方差; h2mg: 主基因遗传率。
m: Total average; da: Additive effect for the first major gene; db: Additive effect for the second major gene; ha: Dominance effect for the first major gene; hb: Dominance effect for the second major gene; i: Additive-by-additive for the two major genes; Jab: Additive-by-dominance for the two major genes; jba: Dominance-by-additive for the two major genes; l: Dominance-by-dominance interaction effects for the two major genes; σ2mg: Genetic variance major-gene; h2mg: Heritability for major-gene.
3 讨论
青海主栽春小麦品种‘青春38[21]由青海省农林科学院于1975年以‘Consens(加拿大红麦)为母本,以冬麦‘03702号בW97208杂交后代为父本杂交选育而成,具有抗倒伏,耐寒,耐青干,高抗条锈、叶锈、秆锈等特点。本研究组从2010年-2016年连续大田成株期抗条锈性观察,发现‘青春38一直表现高抗,由于两个试验地至2016年为止主要流行的条锈病生理小种为CYR32和CYR33(数据未发表),‘青春38成株期对我国条锈病主要流行小种CYR32和CYR33表现为高抗。
国内目前对小麦成株期抗条锈病基因遗传分析的报道主要是根据孟德尔遗传规律进行分析,获得1对、2对或3对抗条锈病基因的作用规律[22-27]。本研究中‘青春38与‘Taichung 29杂交群体后代的抗病性结果不符合纯数量性状遗传的连续性分布,而抗病性鉴定结果中抗病反应中间型较多,进行抗病、感病劃分时误差较大,特别是对严重度结果的划分比较困难,难以根据孟德尔经典遗传规律进行简单的抗病、感病单株(或家系)数目分离比分析,故而应用植物数量性状主基因+多基因混合遗传模型单个分离世代分析方法分析‘青春38成株期抗病基因的遗传特点和作用方式。
本研究选取春小麦品种‘青春38和‘Taichung 29及其杂交后代,分别在西宁和互助两个地区进行田间条锈病抗性鉴定试验。结果表明,‘青春38在成株期的抗条锈性是由2对主基因控制,只是在不同环境下,或是用不同参数(严重度或反应型)表示的情况下,主基因的作用方式有所变化。根据‘青春38的抗性遗传特点,可通过抗病基因聚合以进一步提高寄主的抗病性和抗性持久性,也可以进一步定位抗病基因,获得与抗病基因紧密连锁的分子标记,为分子标记辅助选择育种服务。
参考文献
[1] CHEN Xianming. Epidemiology and control of stripe rust (Puccinia striiformis f.sp. tritici) on wheat [J]. Canadian Journal of Plant Pathology, 2005, 27: 314-337.
[2] 陈万权, 康振生, 马占鸿, 等. 中国小麦条锈病综合治理理论与实践[J]. 中国农业科学, 2013, 46(20): 4254-4262.
[3] LINE R F, CHEN Xianming. Successes in breeding for and managing durable resistance to wheat rusts [J]. Plant Disease, 1995, 79: 1254-1255.
[4] 姚强,郭青云,闫佳会,等.青海东部麦区小麦条锈菌越冬调查初报[J].植物保护学报,2014,41(5):578-583.
[5] 王建超,冯晶,王凤涛,等.我国小麦农家品种‘小红芒成株抗条锈性遗传分析[J].植物保护,2015,41(1):154-157.
[6] 刘太国,邱军,周益林,等.中国冬小麦区域试验品种抗病性评价[J].中国农业科学,2015,48(15):2967-2975.
[7] 王吐虹,郭青云,蔺瑞明,等.中国40个小麦农家品种和甘肃南部40个生产品种抗条锈病基因推导[J].中国农业科学,2015,48(19):3834-3847.
[8] 王步云,冯晶,王凤涛,等.河南省小麦品种(系)遗传多样性和抗条锈性分析[J].华北农学报,2014,29(S1):62-70.
[9] 高晓梅,冯晶,蔺瑞明,等.2008-2009年度河北、河南、四川和江苏四省冬小麦区试品系的遗传多样性和抗条锈性分析[J].植物保护,2014,40(2):113-118.
[10] 王步云,冯晶,蔺瑞明,等.2007—2008年度冀鲁豫川四省小麦区试品种(系)遗传多样性和抗条锈性分析[J].中国农学通报,2014,30(28):255-263.
[11] 曹世勤,吕小欢,黄瑾,等.甘肃省26个春小麦品种(系)苗期抗条锈基因分析及成株期抗病性评价[J].麦类作物学报,2013,33(4):771-776.
[12] 曹世勤,王晓明,贾秋珍,等.2003-2013年小麦品种(系)抗条锈性鉴定及评价[J].植物遗传资源学报,2017,18(2):253-260.
[13] 李永平,曹世勤,金社林,等.临麦系列春小麦品种抗条锈性分析[J].植物保护,2016,42(2):209-213.
[14] 王万军,曹世勤,王晓明,等.86份贵协系小麦种质资源对条锈病的抗病性评价[J].植物保护,2016,42(2):198-203.
[15] 周新力,詹刚明,黄丽丽,等.80份国外春小麦种质资源抗条锈性评价[J].中国农业科学,2015,48(8):1518-1526.
[16] 姚强,贺苗苗,闫佳会,等.春小麦品种青春39的抗条锈性遗传分析[J].麦类作物学报,2014,34(1):39-42.
[17] 姚强.120个春小麦品种(系)成株期抗条锈性鉴定与评价[J].西北农业学报,2014,23(7):96-101.
[18] 侯璐,闫佳会,姚强,等.81份春小麦种质资源抗条锈病评价[J].植物保护,2017,43(3):171-176.
[19] 侯璐.4个春小麦种质资源抗条锈性鉴定和抗性遗传分析[J].麦类作物学报,2017,37(1):80-86.
[20] 刘兵.植物数量性状分离分析Windows软件包SEA的研制[D].南京:南京农业大学,2013.
[21] 刘青元.青海省农作物品种志(1983-2005)[M].西宁:青海人民出版社,2008:51.
[22] 张莹,周新力,王琪琳,等.小麦品系P9897成株期抗条锈性遗传分析[J].麦类作物学报,2015,35(10):1355-1359.
[23] 周春宏,徐智斌,冯波,等.西藏地方小麦品种曲白春的抗条锈性遗传分析[J].麦类作物学报,2015,35(3):312-316.
[24] 马东方,周新力,井金学,等.小偃6号成株期高温抗条锈性遗传分析[J].植物保護学报,2013,40(1):33-37.
[25] 宁利园, 郄彦敏, 王凤涛, 等. 中国小麦农家品种红锁条和白蚂蚱的抗条锈性遗传分析[J]. 植物保护学报, 2015, 42(2): 145-152.
[26] 苏萍萍,曾庆东,李海洋,等.小麦种质P10078的成株期条锈病抗性特征及遗传规律[J].麦类作物学报,2017,37(4):500-503.
[27] 李邦发.西科麦2028抗条锈性的遗传分析[J].植物遗传资源学报,2015,16(5):1093-1097.
(责任编辑: 杨明丽)
