旋毛虫副肌球蛋白在BaculoDirect昆虫杆状病毒表达系统中的表达及鉴定*
2018-11-10王子霞郝春悦黄京京程喻力诸欣平
王子霞 郝春悦 赵 夕 黄京京 程喻力 诸欣平
(首都医科大学基础医学院病原生物学系,北京 100069)
旋毛虫病是一种呈世界性广泛分布的食源性人兽共患寄生虫病。该病严重影响人体健康及畜牧业生产,阻止家畜尤其是猪的感染是减少人类患病的重要措施。迄今,尚无有效的兽用抗旋毛虫疫苗问世(Baietal., 2017)。制备有效的旋毛虫抗原是研制疫苗的关键步骤。本课题组前期研究首次发现并克隆了旋毛虫副肌球蛋白(TsPmy,GenBank登陆号为EF429310),并证实重组蛋白(rTspmy)具有较好的免疫学活性和免疫保护性,以及免疫调节功能(Sunetal., 2015)。然而,由于前期对TsPmy的研究基于原核表达系统,重组蛋白表达为包涵体形式,即无活性蛋白聚集体、复性困难,且原核表达的宿主E.coli翻译后修饰功能有限,不能产生N-和O-端糖基化、脂肪酸酰化、磷酸化以及二硫键等修饰(Dalyetal., 2005),而这些修饰对于活性蛋白的生物活性、功能、结构、溶解度等都非常重要,因此限制了对TsPmy功能的进一步研究(范翠英等, 2012)。
目前,常见的真核表达系统有酵母表达系统、昆虫杆状病毒表达系统、哺乳动物细胞表达系统。酵母表达系统简单高效,有的蛋白可分泌至细胞外,易于纯化,但是该系统蛋白产物不均一,具有密码子偏好,一个基因能否成功表达具有较大随机性;哺乳动物表达系统成本高、操作复杂、产量低;而昆虫杆状病毒表达系统以杆状病毒作为外源基因载体,具备翻译后修饰功能,例如正确的蛋白折叠、寡聚、磷酸化、糖基化、酰基化、二硫键的形成、蛋白水解剪切等,可提高蛋白可溶性,这一系列的特性使其具备成功表达复杂结构蛋白的可能。另外,由于杆状病毒的宿主是无脊椎动物昆虫,不会感染其他动物、植物、人类,故认为杆状病毒的应用较为安全,因而被广泛地应用于基因工程、药物开发、疫苗生产、免疫活性分子和某些致瘤病毒蛋白的表达以及基因表达调控的研究等多个领域中,其表达的重组蛋白可具有天然免疫功能(Smithetal., 1983)。迄今为止,已有上千基因在昆虫细胞或幼虫体内得到高效表达,为获得大量的类原型蛋白及其功能研究提供了可行性(van Oersetal., 2015)。本研究拟利用昆虫杆状病毒表达系统表达rTsPmy,以期获得生物学活性较好的重组蛋白,为旋毛虫病疫苗的研制奠定基础。
1 材料与方法
1.1 材料
1.1.1杆状病毒、细菌、细胞:Sf9昆虫细胞购自Gibco公司,杆状病毒购自Invitrogen公司,感受态DH5α菌购自北京天根生化科技公司,感受态DB3.1菌(质粒pDONR221)以及大肠杆菌BL21(DE3)菌株来自本实验室保存。
1.1.2主要试剂:TsPmy/pET28 a(含TsPmy基因)由本实验室构建并保存;Gateway BP ClonaseII Enzyme Mix and Reagents、BaculoDirect N-Term Transfection Kit、更昔洛韦(gancicovir)购自Invitrogen公司,SFX-Insect Media、胎牛血清(FBS)购自Hyclone公司,Sf-900 III SFM、Grace′s Medium, unsupplemented、0.25%胰蛋白酶(含0.25%EDTA)购自Gibco公司;双抗(青霉素、链霉素)购自Sigma公司;小鼠抗TsPmy羧基端单克隆抗体(9G3)(Haoetal., 2014)、人工感染旋毛虫小鼠血清以及人工感染旋毛虫猪血清由本实验室制备;His单克隆抗体购自Gene公司;IRDye 800CW标记的羊抗小鼠以及羊抗猪IgG多抗购自LI-COR公司;His Bind试剂盒、Protein Refolding试剂盒购自Novagen公司。
1.2 方法
1.2.1目的基因attB-Pmy的扩增:首先根据GenBankTsPmy基因的编码区全序列2 655 bp(75~2 729 bp)设计含attB位点的TsPmy引物,(上游引物P15′-G G G G A C A A G T T T G T A C A A A A A A G C A G G C T T C A T G T C T C T G T A T C G C A G TC-3′,下游引物P2 5′-G G G G A C C A C T T T G T A C A A G A A A G C T G G G T C C T A A T A T T C A T G T C C T T C T TC-3′),由上海Invitrogen公司合成。扩增条件:98 ℃ 5 s;94 ℃ 15 s,55 ℃ 30 s,72 ℃ 3 min,30个循环;最后72 ℃ 3 min。PCR产物进行0.8 %琼脂糖凝胶电泳初步鉴定。
1.2.2重组杆状病毒的构建:利用试剂盒将纯化后目的片段与供体载体质粒pDONR221(含attP位点)进行Gateway BP重组反应构建入门载体,反应条件25 ℃孵育1 h。产物转化感受态大肠杆菌DH5α,转化产物涂在含卡那霉素的LB固体培养基平板进行筛选,挑取阳性菌落进行PCR鉴定:95 ℃ 5 s;94 ℃ 15 s,55 ℃ 30 s,72 ℃ 3 min,30个循环;最后72 ℃ 3 min。将PCR鉴定阳性结果送至上海Invitrogen公司测序,选取测序正确的质粒作为入门载体质粒(pDONR221/Ts-Pmy),利用BaculoDirect N-Term Transfection Kit,通过BaculoDirect线性DNA与pDONR221/Ts-Pmy进行Gateway LR重组反应,构建重组杆状病毒DNA(含his tag),反应条件为25 ℃,18 h。
1.2.3重组杆状病毒转染Sf9细胞:取悬浮培养对数生长期(1.5~2.5×106cells/mL)的Sf9昆虫细胞接种至6孔板(8×105个/孔),培养基为Grace′s Insect Medium,Unsupplemented,27 ℃过夜培养使其贴壁生长,利用转染试剂CellfectinⅡ Reagent、LR重组反应液进行转染实验。将重组杆状病毒DNA导入Sf9细胞,同时加入更昔洛韦100 μmol/L进行药物筛选:将六孔板置于湿润、密封、避光的无菌盒内,27 ℃孵育7 d左右,直到出现病毒感染现象,此时收集培养上清液,作为第1代重组病毒P1,经所获病毒液反复感染Sf9细胞,以获得滴度较高的重组病毒(P3,P4)。
1.2.4重组蛋白的表达、纯化及免疫学鉴定: 悬浮培养Sf9细胞(2~4×106cells/mL),以较高滴度重组病毒感染Sf9细胞。由于rTspmy为胞内表达,不分泌至细胞培养上清,且BaculoDirect线性DNA在外源基因插入位点后有6个连续的his-tag序列,因此,分别于感染后0、24、48、72 h收集细胞沉淀,通过超声破碎法制备细胞裂解液,以低温高速离心(4 ℃, 10 000×g)分离沉淀和上清,应用SDS-PAGE及Western blotting进行表达时相、表达量以及可溶性分析,其中Western blotting一抗为his tag单克隆抗体(1∶5000)。大量悬浮培养Sf9细胞,于重组病毒感染后96 h收集细胞沉淀,通过超声破碎细胞,采用His Bind试剂盒、Protein Refolding试剂盒对rTsPmy进行纯化、复性,步骤参考试剂盒说明书。使用抗旋毛虫单克隆抗体9G3(1∶20 000)、人工感染旋毛虫小鼠血清(1∶2 000)以及人工感染旋毛虫猪血清(1∶2 000)分别作为一抗,对纯化后的rTsPmy进行免疫学鉴定。
2 结果
2.1 TsPmy重组杆状病毒的构建与表达
2.1.1AttB位点连接的TsPmy基因产物(AttB-Pmy)构建成功:TsPmy基因编码区全序列 2 655 bp,引入attB位点后,大小为2 718 bp,为下一步构建入门载体做准备。P1、P2引物扩增出的片段大小与目的片段长度吻合(图1)。
图1 Pmy基因PCR扩增Fig.1 The amplification of TsPmy gene by PCRM: DNA分子量标准;1: TsPmy基因PCR产物;2: 阴性对照(H2O作为模板)。M: DNA marker; 1: PCR product of TsPmy gene; 2: Negative control (template was H2O).
2.1.2入门载体质粒(pDONR221/TsPmy)鉴定以及测序: BP重组反应构建入门载体质粒(pDONR221/TsPmy),转化之后,挑取pDONR221/TsPmy的阳性菌落,以TsPmy的引物P1、P2进行菌落PCR,阳性菌落扩增片段大小约2.7 kb,扩增后测序结果显示均含有阳性重组质粒pDONR221/TsPmy,重组质粒序列经BLAST比对,显示完全匹配,没有突变或缺失。通过LR反应构建重组杆状病毒DNA。
2.1.3重组杆状病毒DNA鉴定: 以重组杆状病毒DNA为模板,利用杆状病毒线性DNA多角体基因部分为上游引物,TsPmy 3′端作为下游引物,进行PCR,并对产物进行琼脂糖电泳。结果显示扩增片段位于3 000 bp左右,与预期值相符(图2)。表明TsPmy基因已转座到BaculoDirect线性DNA上,得到了重组病毒DNA。
图2 重组病毒DNA的PCR鉴定Fig.2 Identification of recombinant viral DNA by PCRM: DNA分子量标准;1: TsPmy特异性引物鉴定的PCR产物;2:阴性对照(H2O作为模板)。M: DNA marker; 1: PCR product of recombinant viral DNA with TsPmy specific primer; 2: Negative control (template was H2O).
2.2 重组病毒感染昆虫细胞Sf9
以P4重组病毒感染Sf9,感染后每隔24 h在光学显微镜下观察细胞的形态。如图3所示,于转染后48 h细胞出现病毒感染的病理征象:细胞核变圆、变大,细胞体积增大,胞质内出现囊泡、颗粒,增殖速度减慢;72 h大多数细胞已停止增殖,96 h细胞开始裂解(图3)。在72~96 h收集细胞沉淀,以获得重组蛋白;在96 h后或细胞裂解后收集培养上清,以获得重组病毒。
2.3 rTsPmy的表达时相分析及Western blotting鉴定
BaculoDirect线性DNA在rTsPmy基因插入位点后有6个连续的his-tag序列,重组病毒感染Sf9细胞,rTsPmy为胞内表达,不分泌至细胞培养上清。本实验分别取24、48、72 h病毒感染Sf9细胞,经SDS-PAGE电泳、考马斯亮兰染色以及Western blotting分析(His单抗作为一抗),结果显示,感染后24 h,rTsPmy尚未表达;48 h,在全细胞裂解液沉淀及上清rTsPmy均有表达,可见此时rTsPmy呈现部分可溶性表达,可溶表达量较少;72 h时,rTsPmy表达量明显增加,但全细胞裂解液上清中未见目的条带,可见此时rTsPmy已经基本呈不可溶表达,目的条带为110 kDa处特异条带,与插入片段编码蛋白质的理论值相符,未感染的Sf9细胞在相应处无特异条带(图4)。

图3 重组病毒感染的昆虫细胞Sf9形态学变化(400×)Fig.3 Morphological changes of recombinant virus-infected Sf9 cellsA. 未感染重组病毒的Sf9细胞在0、24、48、72 h的状态;B. 重组病毒感染的Sf9细胞在0、24、48、72 h的状态。Sf9细胞为悬浮培养,图中箭头所指为出现囊泡、颗粒的细胞。Uninfected Sf9 cells (A) and infected Sf9 cells (B) in 0,24,48,72 h. Sf9 cells were suspended in culture medium. Arrows refer to the Sf9 cells with vesicles and granules.
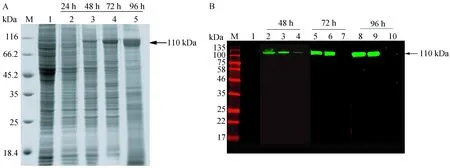
图4 TsPmy的表达时相分析及Western blotting鉴定Fig.4 SDS-PAGE and Western blotting analysis of expression of rTsPmyA. 用SDS-PAGE的方法分析rTsPmy的表达时相。M: 蛋白分子量标准;1: 未感染Sf9全细胞裂解液(1%SDS)作为阴性对照;2-5: 分别为Sf9细胞感染重组病毒24、48、72、96 h后全细胞裂解液(1%SDS)。B. 用Western blotting的方法分析rTsPmy的表达,一抗为his tag单克隆抗体,二抗IRDye 800CW标记的羊抗小鼠多克隆抗体。M: 蛋白分子量标准;1: 未感染Sf9细胞全细胞裂解液(1%SDS);2-4: 分别为Sf9细胞感染重组病毒48 h后全细胞裂解液(1%SDS)、细胞裂解液沉淀(1%SDS)、细胞裂解液上清(PBS);5-7: 分别为Sf9细胞感染重组病毒72 h后全细胞裂解液(1%SDS)、细胞裂解液沉淀(1%SDS)、细胞裂解液上清(PBS);8-9: 分别为Sf9细胞感染重组病毒96 h后全细胞裂解液(1%SDS)、细胞裂解液沉淀(1%SDS)、细胞裂解液上清(PBS)。A. SDS-PAGE analysis of Sf9 cell lysate. M: Molecular markers; 1: Uninfected Sf9 cell lysate (1%SDS) as a negative control; 2-5: Infected Sf9 cell lysate harvested at 24, 48, 72, 96 h after infection (1%SDS). B. Western blotting analysis of Sf9 cell lysate. Samples were detected by anti-his tag monoclonal antibody. The cell lysates were separated as precipitation and supernatant with high speed centrifugation (4 ℃, 10 000×g). M: Molecular size markers; 1:Uninfected Sf9 cell lysate (1%SDS) as negative control; Infected Sf9 cell lysate harvested at 48 h(2-4),72 h(5-7) and 96 h (8-10) after infection respectively (2,5,8 cell lysate (1%SDS); 3,6,9 precipitation (1%SDS); 4,7,10 supernatant (PBS)).
2.4 rTsPmy的纯化及鉴定
由于rTsPmy在昆虫杆状病毒表达系统中可溶表达量较少,因此选择沉淀部分进行his镍柱纯化,并复性。SDS-PAGE电泳后,在约110 kDa处可见单一条带,经his tag单抗以及本实验室制备的9G3单克隆抗体检测,在约110 kDa处可见单一条带(图5)。
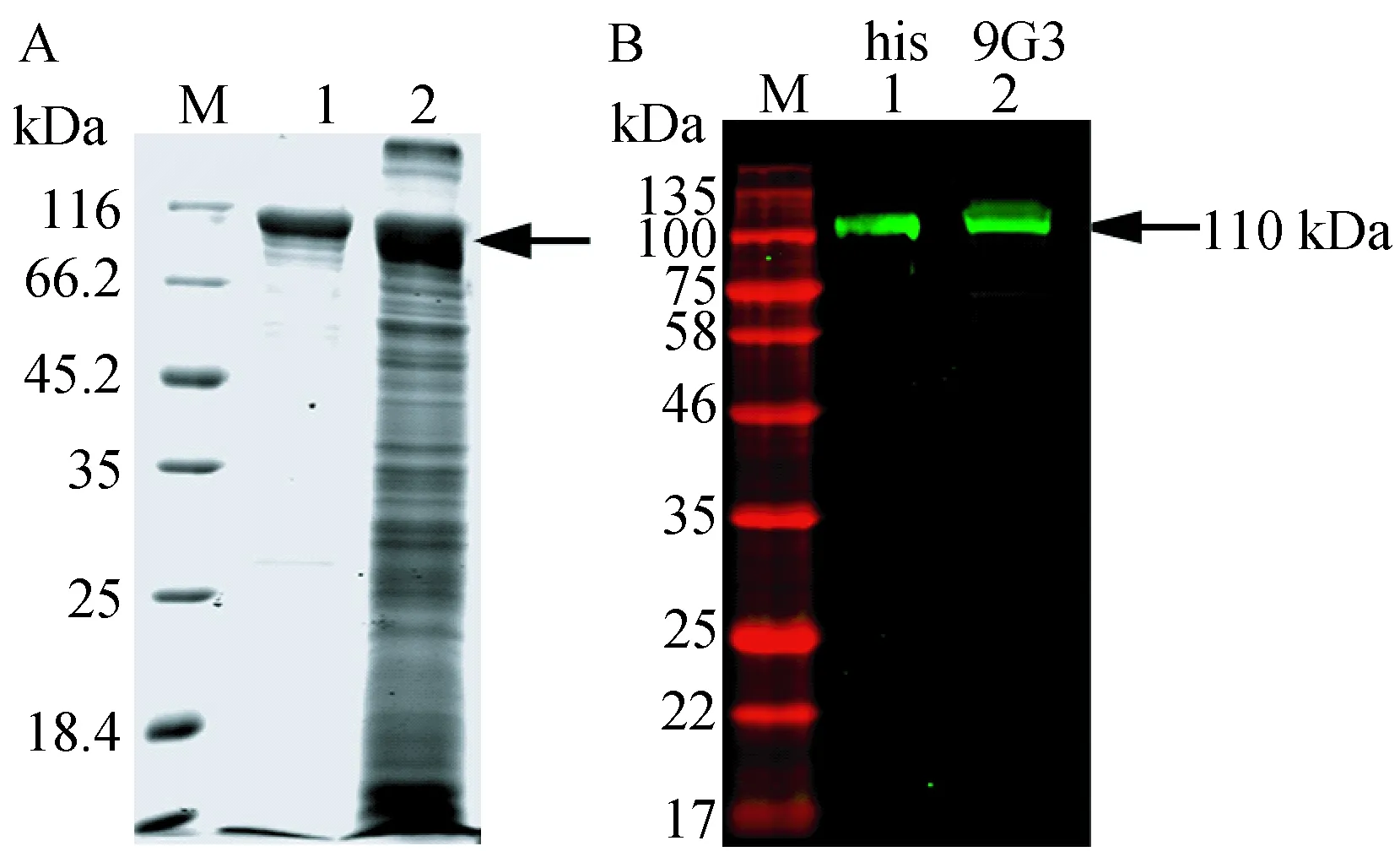
图5 rTsPmy的纯化及鉴定Fig.5 Purification and identification of rTsPmyA. SDS-PAGE检测rTsPmy纯化结果。纯化方法为his镍柱纯化。M: 蛋白分子量标准;1: 纯化rTsPmy; 2: 感染Sf9细胞裂解液。B. 用Western blotting方法鉴定纯化rTsPmy。M: 蛋白分子量标准;1: 一抗为his tag单克隆抗体,二抗IRDye 800CW标记的羊抗小鼠多克隆抗体; 2: 一抗为9G3单克隆抗体,二抗IRDye 800CW标记的羊抗兔多克隆抗体。A. SDS-PAGE analysis of purified rTsPmy. M: Molecular size markers; 1: Purified rTsPmy; 2: Infected Sf9 cell lysate. B. Identification of rTsPmy by Western blotting. M: Molecular size markers; 1: Purified rTsPmy detected by anti-his tag monoclonal antibody; 2: Purified rTsPmy detected by 9G3 monoclonal antibody.
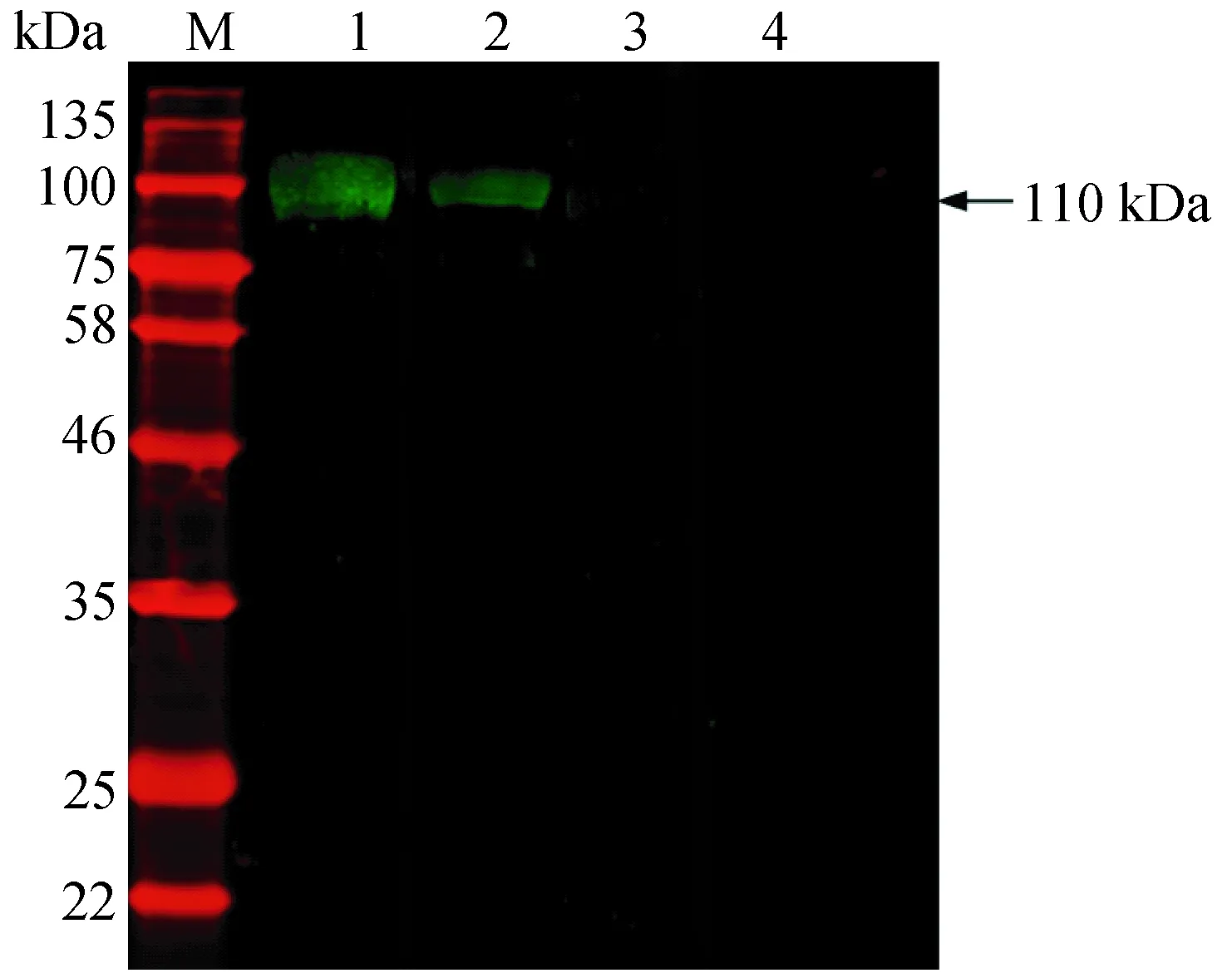
图6 rTsPmy与不同阳性血清的Western blotting鉴定Fig.6 The antigenicity of rTsPmy by Western blottingM: 蛋白分子量标准;1: 人工感染旋毛虫猪血清;2: 人工感染旋毛虫小鼠血清;3:正常猪血清;4: 正常小鼠血清。M: Molecular markers; 1: rTsPmy recognized by infected swine sera; 2: rTsPmy recognized by infected mice sera; 3: rTsPmy recognized by normal swine sera; 4: rTsPmy recognized by normal mice sera.
2.5 rTsPmy免疫学特性鉴定
Western blotting结果显示,rTsPmy能被人工感染旋毛虫的猪抗血清、小鼠抗血清所识别,重组蛋白对阴性对照血清(正常猪血清,正常小鼠血清)均未出现识别信号,表明rTsPmy具有抗原特异性(图6)。
3 讨论
副肌球蛋白是多种无脊椎动物的肌纤维蛋白,例如软体动物、环节动物、节肢动物等,它存在于虫体平滑肌、皮下以及部分寄生虫体表,还可以分泌至体外(Schmitzetal., 1996)。旋毛虫副肌球蛋白有较好的免疫学活性且具有免疫保护作用,在免疫接种小鼠获得36.2%的肌幼虫减虫率,是一个重要的疫苗候选蛋白(Yangetal., 2008)。TsPmy在原核表达系统中出现的不完全表达、纯化效果不佳、有内毒素污染且为包涵体沉淀形式等问题,这些问题的存在限制了对TsPmy功能的深入研究。为了进一步深入研究TsPmy的功能,本实验利用昆虫杆状病毒表达系统成功表达了rTsPmy,且经纯化后能被感染血清有效识别,提示经该表达系统获得的rTsPmy具有较好的抗原性。相对于原核表达系统而言,昆虫杆状表达系统具备明显的优势,包括:该表达系统无内毒素污染,有较完善的转录、翻译后加工修饰功能,使得重组蛋白生物学活性高;表达过程中无血清培养有利于重组蛋白的纯化等。相较于其他真核表达系统而言,它可在多角体启动子控制下高效表达,蛋白产量相对较高(秦顺晴等, 2012)。另外,本实验选用BaculoDirect杆状病毒表达系统,不同于传统的Bac to Bac杆状病毒表达系统,避免了大肠杆菌冗长耗时的定向转座或昆虫细胞中长时间的同源重组,而是使用了快速的Gateway重组反应来获得转染所需的重组杆状病毒,节省了制备重组病毒需要的数天时间,加快了研究进程,而且,Gateway克隆技术十分灵活,方便了将来载体的替换。
应用昆虫杆状病毒表达系统获得rTsPmy,可避免内毒素的污染,且his单抗检测显示单一条带,避免了原核表达系统出现的不完全表达,后期应用亲和层析柱纯化效果较好。但应用昆虫杆状病毒表达系统获得的rTsPmy也存在一定局限性,虽然优化表达温度和时间可以获得一定量的可溶性rTsPmy,但是通过该表达系统获得的rTsPmy主要为不可溶形式,可能是由于副肌球蛋白是多聚体结构蛋白,分子量较大,本身易以不可溶形式存在(Liuetal., 1998)的原因。
本实验应用昆虫杆状病毒表达系统获得了翻译后修饰更加完善、纯化效果较好的rTsPmy,其生物学活性更适于深入研究TsPmy的功能,该研究为制备有效的旋毛虫病疫苗候选抗原分子提供了生物学基础。
