黄土高原草地次生演替过程中微生物群落对植物群落的响应
2018-11-01白丽范席德王洁莹赵发珠薛科社
白丽,范席德,王洁莹,赵发珠,薛科社
西北大学/陕西省地表系统与环境承载力重点实验室,陕西 西安 710127; 西北大学城市与环境学院,陕西 西安 710127
撂荒地植被演替属次生演替,是群落内部关系与外界环境中各种生态因子综合作用的结果(王兴等,2014)。近年来,关于演替动力学的研究一直是植物群落生态学的核心主题(Zhou et al.,2017)。据报道(Zak et al.,2003;胡婵娟等,2012),植被作为研究次生演替动力学的重要组成部分,是生态系统物质循环和能量流动的中枢,而微生物作为地球生态系统中最活跃的组成部分,是生物地球化学循环的主要参与者和重要驱动者,在评价生态系统、维护生态平衡方面起着举足轻重的作用。大量的研究表明,次生演替不可避免地会引起地上植物群落(León et al.,2016;Sun et al.,2017)和地下微生物群落(王艳云等,2016;陈孟立等,2018;Ren et al.,2017;Smith et al.,2015)的变化,这种变化可能对陆地生态系统乃至全球变化有着重要影响。因此,了解植被和微生物在次生演替过程中的动态变化可为陆地生态系统的恢复提供一定的启示(Smith et al.,2015)。
近年来,关于次生演替对植物群落和微生物群落的动态变化已有广泛报道,越来越多的学者开始将研究的热点转移到次生演替过程中植物群落和微生物群落之间的相互作用(Zhang et al.,2009;Tscherko et al.,2004;Han et al.,2007;Lozano et al.,2014)。例如:有研究指出,演替过程中植物多样性和丰富度的增加促使微生物多样性显著增加(Schlatter et al.,2016;He et al.,2008),并且植物均匀度可以通过调节植物物种丰富度从而对微生物群落产生影响(Lamb et al.,2011)。然而,也有一些研究指出,演替过程中植物多样性与微生物群落之间没有直接的相关性(Millard et al.,2010;Han et al.,2007;李毳等,2018);且植物物种丰富度对微生物生物量和多样性具有负面影响。可以看出,植物群落与微生物群落之间存在着复杂的关系,二者之间的相互作用常常出现不一致的研究结果,表明植物群落与微生物群落之间的相互作用依然存在很大的不确定性。这种不确定性对于我们了解植被-土壤系统物质循环、能量流动和信息传递有着极大的阻碍作用。因此,进行演替过程中植物群落与微生物群落之间的相互作用研究显得尤为重要和迫切。
黄土丘陵区是中国严重的水土流失区之一,草地次生演替是防止该区水土流失和恢复该区生态系统的重要管理措施之一。目前,由农田转化为森林的植被次生演替在国内外已经得到了广泛研究,而关于农田转化为草地的相关研究却相对匮乏,这在很大程度上限制了我们对次生演替后植物群落与微生物群落动态变化的全面认识。此外,最近有研究表明,草原系统土壤碳储量和生物多样性均高于森林系统(Zhao et al.,2014;Li et al.,2012)。因此,本研究主要针对黄土高原撂荒若干年后的草地,研究黄土高原草地次生演替20 a、30 a和40 a植物群落和微生物群落特性的动态变化。主要研究目的为:(1)明确草地次生演替过程中植物群落和微生物群落的动态变化;(2)揭示植物群落和微生物群落在早期次生演替过程中的相互作用。
1 研究地区与研究方法
1.1 研究区概况
研究区位于陕西省安塞县纸坊沟流域(109°26′~109°28′E,36°74′~37°77′N)。该区属于典型的黄土丘陵沟壑区,具有典型的温带大陆性半干旱气候。海拔高度为1087.3~1290.8 m,年均温为8.8 ℃,年均降雨量为510 mm左右,大部分降水主要集中在7—9月,占全年降雨量约60%(Zhang et al.,2016)。土壤类型为易受侵蚀的黄绵土(表1)。研究区植物群落的主要物种有铁杆蒿(Artemisia sacrorum)、杠柳(Periploca sepium)、芦苇(Ditch reed)、隐子草(Cleistogenes squarrosa)、苔草(Carex)和大针茅(Stipa grandis)。
1.2 样品采集和分析
于2016年9月,采用时空替代法(Zhang et al.,2016),选取坡向、坡度、海拔高度基本一致的撂荒地作为对照(CK),以自然演替20 a、30 a和40 a的草地作为实验样地,样地基本信息见表2。每个演替年限设置3个重复样方,采用样方法在每个样方中随机选取6个小样方(1 m×1 m)进行植物性状调查,包括植物覆盖度、高度、频度和优势种,对地上部分进行修剪和干燥,获取地上生物量并分析生物量碳。按照Zhang et al.(2016)等文献中描述的方法计算植物多样性、丰富度和均匀度。用直径为5 cm的土钻在每个样方沿S型分别采集表层0~20 cm的土壤样品10个,之后混合为1个土壤样品,过2 mm筛,去除根系和凋落物残体,然后将一部分样品立即运输到实验室并保存在-80 ℃冰箱用于DNA分析;另一部分样品风干处理后用于土壤理化性质测定。
1.3 土壤基本理化性质分析
有机碳、总氮、硝态氮、铵态氮、pH和容重参照鲍士旦(2000)中的方法进行分析测定。
1.4 土壤DNA提取,PCR扩增和高通量测序
采用Omega DNA试剂盒从0.5 g新鲜土壤中提取微生物DNA,使用分光光度计测量总DNA的浓度。使用细菌通用引物 515F(5'-GTGCCAGCMGCCGCGG-3')和 907R(5'-CCGTCAATTCMTTTRAGTTT-3')进行 16S rRNA的PCR扩增(Ren et al.,2017)。使用真菌的通用引物 FR1(5'-AICCATTCAATCGGTAIT-3')和FF390.1(5'-CGWTAACGAACGAGACCT-3')进行18S rRNA的PCR扩增(Cassman et al.,2016)。细菌扩增液的成分组成(Ren et al.,2017):1.25 μL模板DNA(10.0 ng),0.4 μL双链体,0.4 μL Taq聚合酶。PCR循环参数为:95 ℃,5 min;95 ℃,30 s;55 ℃,30 s;72 ℃,60 s;72 ℃,10 min。真菌扩增液的成分组成(王艳云等,2016):1.5 μL模板 DNA(10.0 ng),1.0 μL 引物(50 μmol·L-1),5 μL 10×PCR buffer,0.5 μL dNTP(10 mmol·L-1),0.5 μL Taq聚合酶。PCR循环参数为:95 ℃,3 min;95 ℃,30 s;55 ℃,30 s;72 ℃,30 s;72 ℃,5 min。每个样品扩增3份,然后将对应的扩增产物混合,最后将每个样品等量PCR产物放入管中,送至上海生物研究所于 Illumina MiSeq平台下使用 USEARCH v5.2.32进行测序分析。去除序列中的嵌合体,通过Blast比对数据库对处理后的序列进行物种分类;将相似性大于 97%的序列归为同一种操作分类单元(OTU)(王艳云等,2016)。最后,将完整的数据集发送到国家生物技术信息中心(NCBI)的序列储存(SRA)数据库,序列号为SUB4145523。
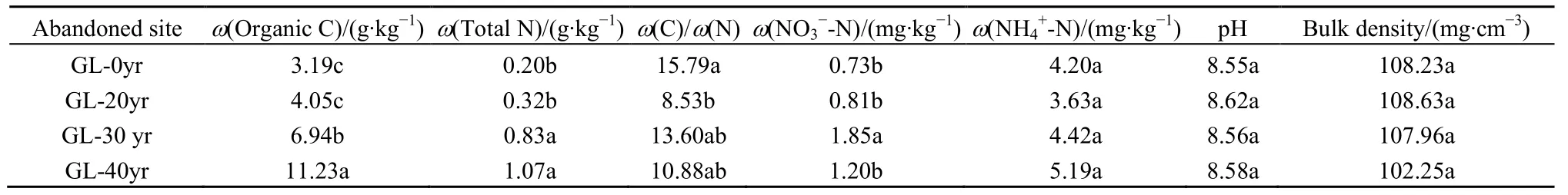
表1 样地土壤基本理化性质(0~20 cm)Table1 Soil physicochemical properties of plots (0~20 cm)

表2 样地植被与地理信息概况Table 2 Status of vegetation and geographical information
1.5 数据统计分析
采用单因素方差分析(SPSS)分析植物群落多样性、丰富度、均匀度、覆盖度、生物量和生物量碳(P<0.001)的差异显著性。采用RDA对植物群落和微生物群落的相关特性进行分析(Canoco 4.5),采用NMDS对微生物β多样性进行分析。文中图示均用Origin 8.5制图。
2 结果分析
2.1 草地演替对植物群落的影响
随着草地次生演替的进行,不同演替阶段植物群落表现出不同的演替趋势(表3)。草地演替20~30年期间,植物覆盖度、丰富度、多样性、均匀度和生物量碳分别增加了2.44%、9.86%、6.86%、5.17%和9.01%,植物生物量基本保持不变;演替30~40 a期间,植物覆盖度、丰富度、多样性、均匀度、生物量和生物量碳分别增加了43.64%、28.53%、6.12%、4.92%、54.55%和 2.16%。植物覆盖度、丰富度、多样性、均匀度、生物量和生物量碳含量在草地演替40 a分别是草地演替20 a的47.13%、41.20%、13.40%、10.34%、54.55%和 11.85%。植物群落在整个演替过程中与植被进展演替方向保持一致,均呈显著增加的趋势(P<0.001)。
2.2 草地演替对微生物群落的影响
2.2.1 草地演替对微生物群落多样性的影响
随着草地次生演替的进行,微生物多样性整体呈现出增加的趋势(图1,图2)。细菌多样性在草地演替20 a、30 a和40 a分别比撂荒地显著增加1.77%、4.17%和4.01%(P<0.01),在演替20~30 a这一阶段增长最快,至演替30 a达到最大,之后出现缓慢下降;真菌多样性与细菌多样性的演替趋势略有不同,与撂荒地相比,真菌多样性在草地演替20 a、30 a和40 a后分别增加了13.79%、23.79%和26.21%,在整个次生演替过程中呈持续增长状态,但不同演替年限间没有表现出显著性差异。

图1 演替对土壤微生物香农多样性的影响Fig. 1 Impact of succession on shannon diversity of soil microbial
2.2.2 草地演替对微生物群落组成的影响
草地次生演替过程中同样伴随着微生物群落组成的变化(图3),如相对丰度>1%的优势细菌门如下:放线菌(Actinobacteria)(34.8%)、变形菌(Proteobacteria)(26.0%)、酸杆菌(Acidobacteria)(15.0%)、绿弯菌(Chloroflexi)(7.5%)、芽单胞菌(Gemmatimonadetes)(8.7%)、硝化螺旋菌(Nitrospirae)(1.6%)、拟杆菌(Bacteroidetes)(2.1%)、疣微菌(Verrucomicrobia)(1.1%)和浮霉菌(Planctomycetes)(1.0%)。其中,放线菌和绿弯菌的相对丰度随草地演替的进行而显著下降,变形菌、酸杆菌、芽单胞菌、硝化螺旋菌、拟杆菌和浮霉菌的相对丰度随草地演替的进行而显著增加(P<0.05),疣微菌随演替进行整体呈增加的趋势,但不显著(表 4)。相对丰度>1%的优势真菌门如下:子囊菌(Ascomycota)(67.19%)、担子菌(Basidiomycota)(16.23%)和接合菌(Zygomycota)(10.43%)。其中,子囊菌和接合菌的相对丰度随演替进行呈增加趋势,担子菌的相对丰度随演替进行呈下降趋势,但都不显著(表5)。
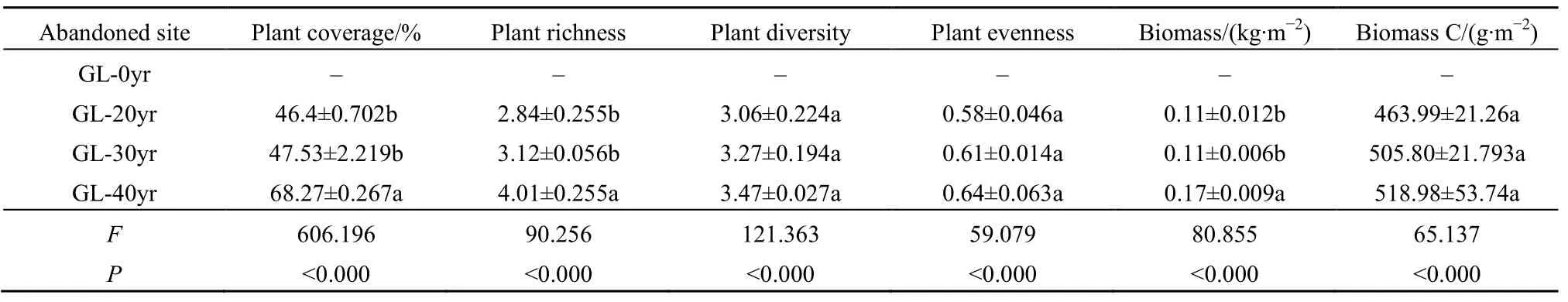
表3 不同演替阶段植物群落特征Table 3 Characteristics of plant communities at different successional sites

图2 草地演替过程中土壤微生物群落比较基于Bray-Curtis差异的NMDS图(A:细菌;B:真菌)Fig. 2 Soil microbial community comparisons by NMDS plots based on Bray-Curtis dissimilarity during succession (A: Bacteria; B: Fungi)

图3 土壤微生物群落在门水平上的相对丰度(A:细菌;B:真菌)Fig 3 Relative abundance of the soil microbial communities at the phylum level (A:Bacteria; B:Fungi)

表4 根据RDP分类研究草地土壤中细菌优势菌群的相对丰度,用广义线性模型评价土地利用对细菌相对丰度的影响Table 4 Relative abundances of dominant bacterial Phylum groups according to the RDP Classifier in soils of studied grassland stands, and effect of land use on relative abundance evaluated by generalized linear model (GLM)
2.3 植物群落和微生物群落之间的关系
基于Pearson相关性分析探讨植物群落与微生物群落多样性的关系(表6),土壤细菌多样性在演替过程中与植物特性(覆盖度、丰富度、均匀度、多样性、生物量和生物量碳)呈显著正相关(P<0.05),土壤真菌多样性与植被特性在演替过程中有相同的变化规律(P<0.05)。RDA分析表明(图4),细菌门类水平中除绿弯菌、拟杆菌和疣微菌菌门相对丰度与植被特性没有呈现出显著相关性之外,其余6个细菌菌门相对丰度与植被特性中的植物覆盖度、生物量、丰富度、多样性、均匀度和生物量碳均呈显著相关(P<0.05)。其中,放线菌与植被特性呈显著负相关,变形菌、酸杆菌、芽单胞菌、硝化螺旋菌和浮霉菌与植被特性呈显著正相关;真菌门类水平中担子菌与植物特性呈显著负相关(P<0.05),子囊菌和接合菌与植被特性无显著相关性。

表6 微生物多样性与植物特性的皮尔森相关系数Table 6 Pearson correlation coefficients between the microbial diversity and the plant traits
3 讨论
3.1 草地演替对植物群落的影响
次生演替是地表同一地段各种不同植物群落按时间顺序分布和替代的过程(梁健,2011)。本研究表明,在整个演替过程中,植物群落物种丰富度、均匀度、覆盖度、多样性、生物量和生物量碳均随演替的进行而显著增加(P<0.001),表明次生演替是影响植物群落特性的主要因素之一(Chabrerie et al.,2003;刘勇,2010)。大多数研究表明,植物丰富度和多样性随演替的推进呈增长趋势(胡婵娟等,2012;王兴等,2014),与本研究结果相一致;然而,刘勇(2010;2007)研究发现随着植被正向演替的进行,植物群落丰富度和多样性明显下降,这可能是因为在群落演替过程中群落内部存在着激烈的生态位竞争。此外,Sun et al.(2017)研究发现,草本植物多样性呈现出驼峰状演替形式,这可能与研究区生境的高度异质性有关(Ruprecht,2005)。总之,次生演替是植物群落对起始阶段异化的过程,不但体现在物种的竞争上,也体现在环境条件的改变上,使生境更适合于演替后来种(黄懿梅,2008)。对次生演替过程中草地植物群落的动态变化进行分析有助于维护和管理生态系统。

表5 根据RDP分类研究草地土壤中真菌优势菌群的相对丰度,用广义线性模型评价土地利用对真菌相对丰度的影响Table 5 Relative abundances of dominant fungal Phylum groups according to the RDP Classifier in soils of studied grassland stands, and effect of land use on relative abundance evaluated by generalized linear model (GLM)

图4 微生物群落组成与植物性状之间关系的冗余分析(RDA)图Fig. 4 Ordination plots of the results from the redundancy analysis to identify the relationships between the abundance of microbial taxa and plant traits
3.2 草地演替对微生物群落的影响
3.2.1 草地演替对微生物群落多样性的影响
次生演替不仅会引起植物群落特性的变化,同时还伴随着微生物群落特性的变化(王静娅等,2016;陈孟立等,2018)。本研究表明,微生物多样性整体随演替时间的增加而增加,这可能是由于演替改善了土壤环境,为微生物的生长提供营养物质,增加了微生物群落的可变性(何寻阳等,2009);而且,在演替初期真菌多样性的增幅高于细菌多样性的增幅,这可能与细菌内部的遗传均一性和真菌内部的遗传变异性有关(Chabrerie et al.,2003)。值得注意的是,我们发现随着演替的进行微生物多样性虽然呈现出增长的趋势,但增长速率逐渐变缓,这是由于随着演替的进行,草地逐渐趋于成熟阶段,而微生物对外界环境变化较敏感(Moghimian et al.,2017;陈孟立等,2018;杨文航等,2017),在一定程度上可以用来指示次生演替的早期动态变化过程(胡婵娟等,2012;Zhang et al.,2009)。
3.2.2 草地演替对微生物群落组成的影响
为了更好地了解微生物群落在次生演替过程中的变化,针对微生物分类群在门类水平上的变化开展了进一步的研究。研究发现,除疣微菌之外,细菌中大多数菌门与植被恢复年限呈显著相关(P<0.05),这可能是由于细菌属r对策物种(Zhou et al.,2017),具有生长速率快、周转率高的特点。同时研究表明,放线菌和绿弯菌在演替初期含量高于演替后期,一方面是由于放线菌和绿弯菌能够抵抗恶劣的生存环境,在养分贫瘠的土壤中具有较强的适应能力(Delgado-Baquerizo et al.,2016);另一方面是由于它们能够较好地控制裸地土壤,促使其含量在演替初期较高(Suela et al.,2013)。变形菌随演替逐渐增加,这是由于变形菌属共营养菌,是恢复土壤养分的功能菌(Zhang et al.,2016;Zhou et al.,2017)。真菌的物种组成在不同演替阶段没有表现出显著相关性,这可能是由于真菌属 k对策物种(Zhou et al.,2017),具有生长速率慢、周转率低的特点。子囊菌和担子菌作为真菌群落的优势菌群,在演替过程中具有不同的演替趋势,子囊菌相对丰度随演替的进行逐渐增加,担子菌相对丰度的演替趋势与之相反。这可能是由于子囊菌和担子菌都是土壤有机质的主要分解者(王艳云等,2016),在一定程度上处于竞争关系,而子囊菌的基因替代速率显著快于担子菌,使得子囊菌较担子菌具有更快的进化速率,在竞争中处于优势(王海英等,2010)。总之,次生演替可以通过资源和栖息地的变化来影响微生物生长的微环境,从而改变微生物群落的多样性和组成(胡婵娟等,2012;Zhao et al.,2014)。
3.3 草地演替过程中植物群落与微生物群落间的响应
植被通过提供碳化合物影响土壤微生物群落的结构和功能,从而影响地上和地下生态系统的生物地球化学循环(Fujimoto et al.,2004;Chen,2016)。研究发现,随着草地演替的进行,土壤微生物多样性与植物群落特性均呈显著正相关(P<0.05),土壤微生物中绝大多数为异养型,表明植物群落是微生物赖以生存的营养物质和能量来源,可以通过增加食物资源的种类和数量来提高微生物多样性(He et al.,2008;王静娅等,2016;张薇等,2005)。植物覆盖度、丰富度和多样性可以通过提高生态系统净初级生产力来加快植物生物量的周转速率及增加根系分泌物的种类,从而为微生物的生长提供较高的动力(王静娅等,2016;Lamb et al.,2011;Bartelt-Ryser et al.,2005;Haichar et al.,2014),增加微生物群落的可变性。植物群落不仅对微生物多样性具有显著影响,而且对微生物群落组成也有很大的影响(何寻阳等,2009)。本研究表明,植物群落特性与微生物群落组成呈显著相关。其中,放线菌和担子菌与植物特性呈显著负相关(P<0.05),变形菌、酸杆菌、芽单胞菌、硝化螺旋菌和浮霉菌与植物特性呈显著正相关(P<0.05)。刘兴等(2015)研究发现,变形菌和浮霉菌在植物群落演替后丰富度逐渐增加,Ren et al.(2016;2017)也观察到造林后硝化螺旋菌的丰富度显著增加,担子菌相对丰度逐渐下降,与本研究结果相一致。由此表明,植物群落与微生物群落组成之间密切相关,植物群落可以通过提供可溶性有机碳快速改变微生物群落的组成(刘兴等,2015)。更重要的是,Tscherko et al.(2004)和 Cassman et al.(2016)研究发现,在草地次生演替过程中,土壤微生物不仅受地上植被的制约,而且一定程度上还受到土壤养分的影响。土壤微生物群落通过控制有机质的分解和养分循环,进而影响地上植物群落的生长和组成,植被与微生物的相互作用决定了生态系统的恢复过程(Han et al.,2007;丁晶等,2017)。未来我们将会增加对土壤质地和养分的调查,以更好地诠释次生演替过程中微生物的动态变化。
4 结论
综上所述,在草地次生演替过程中,植物群落覆盖度、丰富度、均匀度、多样性、生物量和生物量碳均呈不断增加的趋势,尤其是覆盖度、丰富度、多样性和生物量表现最为显著,演替40 a分别比演替20 a增加了47.13%、41.20%、13.40%、和54.55%。微生物多样性在整个次生演替过程中呈持续增长状态,细菌多样性在演替20 a、30 a和40 a分别比撂荒地显著增加了1.77%、4.17%和4.01%;真菌多样性在演替20 a、30 a和40 a分别比撂荒地增加了13.79%、23.79%和 26.21%。本研究发现,在演替过程中,植物群落与微生物群落密切相关,为陆地生态系统地上植物群落和地下微生物群落建立了重要的联系,有助于维护和管理生态系统。
