长期生态学研究和试验示范为高寒草地的适应性管理提供理论与技术支撑
2018-10-25曹广民张法伟李以康杜岩功郭小伟钱大文
曹广民 林 丽 张法伟 李以康 杜岩功 郭小伟 李 茜 钱大文 樊 博
中国科学院西北高原生物研究所 海北高寒草甸生态系统定位站 西宁 810001
高寒草地是青藏高原的主体植被类型,肩负生产、生活、生态的“三生”服务功能。自 20 世纪 70 年代以来,受到气候变化、人类经营理念和经营方式的强烈影响,高寒草地的生产功能被过度使用,生态系统稳定性发生动荡、失衡乃至丧失,形成大面积退化草地。党和国家十分重视青藏高原的生态功能,进入 21 世纪后,国家斥巨资先后实施了三江源退牧还草、三江源国家公园建设和祁连山国家公园试点工作,借以实现高寒草地功能的良性恢复和区域生态环境的综合治理。40 余年来,中国科学院海北高寒草甸生态系统定位站(以下简称“海北站”)高寒草地生态系统对气候变化和人类干扰的响应、适应及其系统演变的生物学过程为主线,先后开展了高寒草地结构、功能及其系统内的物质交换和能量流动、草地生产力提升、高寒草地对气候变化的响应与适应、代表性高寒草地生态系统的承载力、高寒草地演变的环境-生物-土壤-功能联动机制、基于生态过程的高寒草地适应性分区管理技术和典型受损生态系统功能提升关键技术的研发与示范等方面的研究工作,为保障青藏高原生态屏障功能的发挥和区域可持续发展作出了贡献。
1 通过解析高寒草地生态系统结构、生产力分配与种群繁殖模式,首创高寒草地生物学研究,填补了青藏高原相关领域的研究空白,积累了丰厚的有科学价值的数据,对高寒草地生态系统生态过程研究起到了重要的指导作用
自 20 世纪 60 年代起,受“国际生物学规划”(IBP)和“人与生物圈”(MAB)计划的推动,世界各地相继建立了研究各类生态系统结构、功能的长期定位站。海北站建站初期的 10 多年,主要围绕典型高寒草地植物种群的分布特征、群落结构、繁殖特征及其生产力开展工作,为加深高寒草地生态系统的认识,提供了基础数据。研究发现土壤水分条件是决定高寒草甸植物种群分布的主要因素,其中:
(1)以矮嵩草为建群种所形成的草甸是青藏高原分布最广泛的草地类型之一,群落结构比较复杂,种类组成较丰富,约 30 种 /m2,群落总盖度在 85% 左右,主要分布于排水良好的滩地、坡麓及山地半阴半阳坡。群落结构可划分为 2 层。主要伴生种有异针茅(Stipa aliena)、垂穗披碱草(Elymus nutans)、早熟禾(Poa sp.)、麻花艽(Gentiana straminea)、矮火绒草(Leontopodium nanum)、雪白委陵菜(Potentilla nivea)、甘肃棘豆(Oxytropis kansuensis)、鹅绒委陵菜(Potentilla anserina)、异叶米口袋(Gueldenstaedtia diversifolia)、甘肃马先蒿(Pedicularis kansuensis)、苔草(Carex sp.)、多种毛茛(Ranunculus spp.)等。矮嵩草属寒冷中生短根茎地下芽植物,株高 3—10 cm,生长季单株生物量中,地上生物量/地下生物量平均为 0.43[1]。在矮嵩草的生活周期中总繁殖效力平均为 49.63%,其中有性繁殖效力平均为 3.44%,营养繁殖效力为 46.58%,约是有性繁殖效力的 14 倍[2]。矮嵩草种群土壤水势生态位宽度为 0.574 mpa[3]。
(2)小嵩草为建群种所形成的草甸,结构简单,种类组成较少,约 10 种 /m2左右,常见的伴生种有异针茅、美丽风毛菊(S. superba)、垂穗披碱草、矮火绒草、麻花艽、雪白委陵菜、二裂委陵菜(Potentilla bifurca)、高山唐松草(Thalictrum alpinum)、花苜蓿(Trigonella ruthenica)等。片层结构不明显,仅有 1 层,总盖度在 80% 以上,主要分布于山地阳坡和半阳坡。小嵩草属寒冷旱中生短根茎地下芽植物,植株矮小,高 3—5 cm,生长密集容易形成致密的植毡层。小嵩草在生长季单株生物量地上生物量/地下生物量平均比值为 0.66[1]。在整个生长季节中总繁殖效力平均为 39.67%,其中营养繁殖效力平均为 36.10%,有性繁殖效力平均为 4.11%,营养繁殖效力是有性繁殖效力的 8 倍多[4]。小嵩草种群土壤水势生态位宽度为 0.283 mpa[3]。
(3)藏嵩草为建群种的沼泽化草甸,主要由湿中生多年生草本植物组成,群落结构比较复杂,一般存在 2 个片层结构,组成种类较少,10 余种 /m2,伴生种主要有华扁穗草(Blysmus sinocompressus )、黑褐苔草(Carex atrofusca)、青藏苔草(C.moorcroftii)、星状风毛菊(Saussurea stellla)、重冠紫菀(Aster diplostephioides)、线叶龙胆(Getiana farreri)等。藏嵩草甸草层平均高 15—25 cm,总盖度达 90%—95%。生长季单株生物量中,地上生物量/地下生物量平均比值为 1.49,主要分布于河谷两岸较湿的低阶地和河漫滩生境中[1]。整个生活周期的平均繁殖效力为 22.39%,其中营养繁殖效力平均为 19.46%,占总繁殖效力的 82.87%,而有性繁殖效力平均值只有 2.93%[5]。藏嵩草种群土壤水势生态位宽度为 0.267 mpa[3]。
(4)垂穗披碱草为建群种的群落结构较嵩草草甸复杂,组成种类较丰富,总盖度在 60%—80%,垂穗披碱草植株高 20—60 cm,叶层高 15 cm 左右,一般位居群落上片层结构,残存的矮嵩草位居群落下片层结构。其他伴生种类主要有西伯利亚蓼(Polygonum sibiricnm)、鹅绒委陵菜(Potentilla anserina)、二裂委陵菜、细裂亚菊(Ajania tenuifolia)、甘肃马先蒿、雪白委陵菜、大通獐牙菜(Swertia przewalskii)、雅毛茛(Ranunculus puchellus)等。垂穗披碱草地上生物量远远大于地下部分,二者的平均比值为 3.67,在整个生活周期中营养繁殖效力和有性繁殖效力相差无几,营养繁殖效力平均为 16.25%,有性繁殖效力平均为 15.76%,二者比值只有 1.03。垂穗披碱草种群土壤水势生态位宽度为 0.633 mpa,适于生长在比较温暖向阳的河谷阶地、山麓和土壤疏松的滩地,是原生植被破坏后的次生植被类型。在嵩草草甸内鼠类破坏的裸露区域分布较多,这些区域土壤疏松,水分适中、土壤通气性良好,是垂穗披碱草生长的良好环境。
(5)典型高寒草甸草地一般存在双片层群落结构,位于上层的禾本科牧草。在超载放牧下,由于家畜的选择性啃食,种子繁殖受到抑制,群落构型与繁殖策略发生改变,禾草减少,以根茎繁殖为主的嵩草属植物成为优势种群。植物根系的快速和过度发育,导致根土比和根冠比的增大,加速了地表土壤生物结皮和草毡表层的极度发育,造成土壤水分入渗速率的降低和根系对有限养分的生物固定,致使高寒草甸的退化演替过程依次经历:禾草-矮嵩草群落、矮嵩草群落、小嵩草群落和杂类草次生裸地阶段的退化演替过程,根系环境的暖干化及有限养分的供求失衡是导致高寒矮嵩草最终退化的根本原因[6]。小嵩草草甸草毡表层的极度加厚、开裂与剥蚀,形成大面积剥蚀坑或杂类草-“黑土型”次生裸地,是高寒草甸退化的主要景观特征[7],此种景观占据高寒草甸面积的 60% 左右[8]。由于草毡表层阻止水分入渗[9],使大量小嵩草死亡,随后发生草毡表层剥蚀现象,最终演变为“杂类草次生裸地”(图 1)。
(6)鼠类活动挖掘引起草地的破坏是草地退化的伴生表象,只有在草地退化到一定程度,鼠类活动才会大爆发。鼠类活动的加剧,加速了老化草毡表层的破解,增加了土壤有机物质的矿化速率;鼠丘土壤的速效氮、磷和钾养分相较原生草地分别提高了 54.81%、49.18% 和 7.2%,为植被的再生创造了条件,鼠丘成为植被再生的良好基质[10]。鼠类活动的大爆发可能是自然状况下,草地稳定性自我调节或恢复的一种机制,鼠类是退化草地再造的工程师[11]。该研究表明高寒草甸系统退化过程中系统构件变化特征互为因果,其演替的动力、机制及其稳定性维持的瓶颈各异,基于草地生态过程的适应性管理,是高寒草地可持续利用的关键,为高寒草地的适应性管理提供了理论支撑。
该方面的研究开创了高寒草地过程研究的先河,对诸如高寒草地与全球变化,人类干扰下的高寒草地演变,以及放牧高寒草地的适应性管理及其功能提升技术的选择,提供了丰厚的本底基础数据积累,对当前的研究仍具有重要的借鉴与指导价值。
2 长期定位监测与模拟实验研究,发现高寒草地通过改变生态系统的群落结构,被动选择适宜性生态演化路径的调节策略,减弱了生态系统整体对外界干扰的应激反应,保障了系统的稳定性
青藏高原正在经历着以增温为主要特征的气候变化过程。海北站自 2000 年以来,相继建立了包括模拟增温放牧、模拟水热改变(图 2)、山体垂直带(3 200—3 800 m)双向移栽环境置换、高寒湿地旱化和养分添加等长期实验。发现了一些高寒草地应对环境变化的特殊生物学机制,以及系统稳定性维持的生态过程。
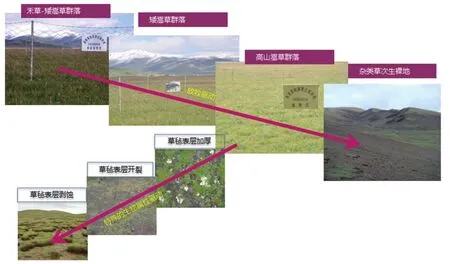
图1 高寒草甸退化过程
海北站连续 32 年的气象观测发现,1980—2014 年海北站地区温度变率为 0.42℃/10 a(p < 0.001),降水变率为 –32.9 mm/10 a(p=0.10)。土壤含水量亦呈线性下降趋势,植物生长季土壤 5 cm 和 45 cm 平均湿度分别由 2002 年、2003 年的 42.7%±5.9% 和 44.2%±2.4% 下降到 2009 年的 24.9%±3.8% 和 31.9%±10.5%,下降了 17.8 和 12 .3个百分比[12]。区域气候呈现暖干化的变化趋势。
草地生产力监测表明,高寒草甸地上初级生产力和高寒灌丛总初级生产力分别为 164.10±4.27 g C m−2a−1和511.8±11.3 g C m−2a−1,其年际变异 2%—5%,草地生产力对气候变化具有较高的稳定性[12,13]。
涡度相关测定数据也表明,高寒嵩草草甸和高寒灌丛草甸生态系统表现出碳的弱汇特征,其值为113.65±93.33 g C m−2a−1和 −74.4±12.7 g C m−2a−1[13,14]。其值大小与草地地上初级生产力基本相当,当草地处于放牧状态下,这部分碳将随食物链的运移最终返回到大气系统,生态系统碳收支基本处于平衡状态。
青藏高原高寒草地碳库容量巨大,其草地总面积 160 万平方公里,总碳储量 26.47 Pg C,占中国植被碳储的 5.1%、土壤碳的 24.3%[15]。高寒草地具有巨大的碳增储潜力,其潜力来源于退化草地恢复对系统植被-土壤碳库的重建。我国北方实施退牧还草和退耕还草,其土壤有机碳固持量 0—40 cm 和 0—30 cm 分别为 130.4 g C m−2a−1和 128.0 g C m−2a−1[16]。但由于青藏高原气候严酷,生态系统脆弱而敏感,人类干扰强度较大,其碳增储潜力的发挥漫长而艰难。
水热改变虽然对草地初级生产力没有造成影响,但改变了高寒草地群落的生物多样性与结构,高寒草地植物群落优势种群发生了由表聚根性的莎草科植物向深根型禾本科植物的转变。植物群落根冠比降低,即深根系的禾草增加、浅根系的莎草减少,Meta 分析也证实了气候造成的这种变化在整个高原普遍存在(图 3)[17]。放牧干扰实验亦发现,随着放牧强度的增加,草地植物群落优势种也会发生这种改变[18,19]。这种功能群组成的变化增强了植物群落对深层土壤水分的获取能力,保障了草地系统的稳定。
山体垂直带双向移栽试验证明,增温使得牧草的3种早期物候事件(返青期、现蕾期和初花期)和晚期物候事件(初黄期和枯黄期)分别提前 4.8—8.2 天 /℃ 和 3.2—7.1 天 /℃;而降温使得 3 个早期物候推迟 3.8—6.9 天 /℃ 和 2 个晚期物候提前了 3.2—8.1 天 /℃,而初果期和终果期的变化则相对较小,以便最大限度地保证果实成熟和扩散,通过保持结果期的相对稳定来适应气候变化[20]。

图2 水热改变对高寒草地影响模拟
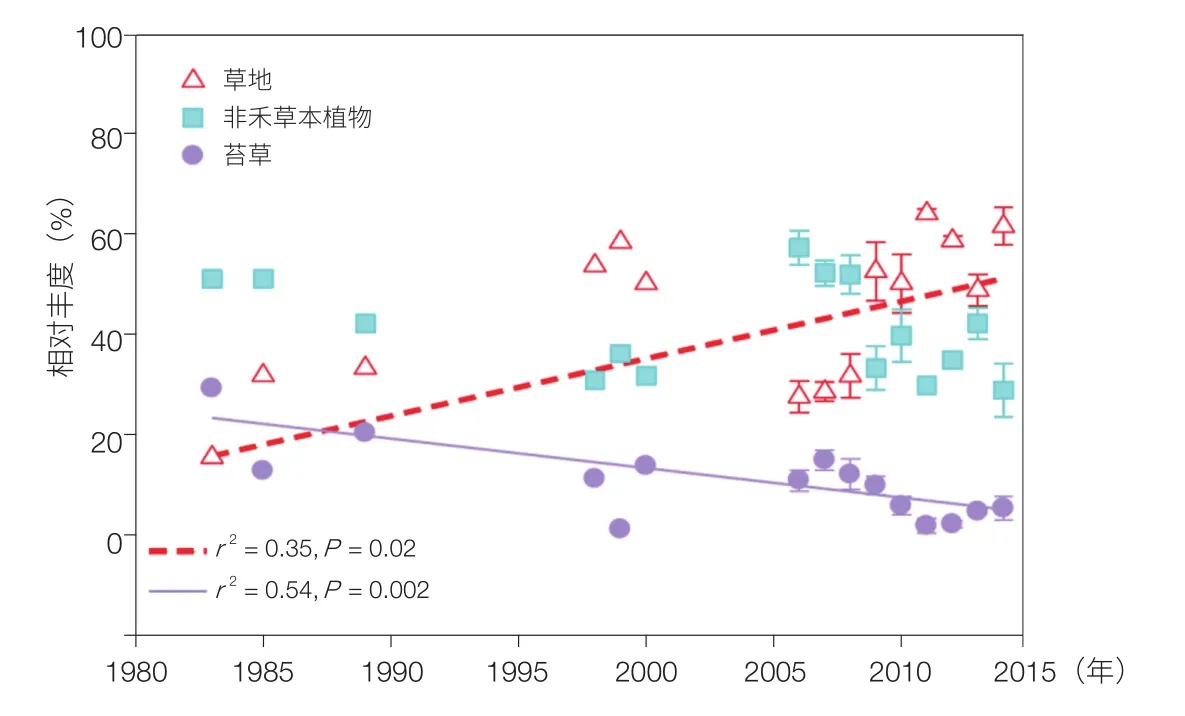
图3 高寒草甸植物功能群的演变
宏基因组学分析表明,湿地旱化和氮沉降可引起 CH4产生潜势的降低,从而导致系统净 CH4排放的减少,硝化潜势的降低和反硝化潜势的增加,共同影响了系统 N2O 的排放。综合 CO2、N2O 和 CH43 种温室气体对旱化的响应,水位降低和氮输入增加均降低了高寒湿地温室气体排放的全球变暖潜势[21]。同时,通常认为,湿地水位下降将通过增强土壤酚氧化酶的活性而促进有机碳的降解。我们研究发现,随着湿地水位下降,伴随着亚铁向铁氧化物的转化,有更多的木质素受到铁氧化物的保护,土壤亚铁的氧化为核心的“铁门”(iron gate)效应,抑制了湿地的碳损失[22]。这种微生物和土壤化学机制,为解释和预测湿地干旱过程中的土壤碳动态提供了新的思路和依据。
高寒环境有利于土壤有机物质的积累,形成草地“全量养分丰富、速效养分贫乏”的特性。高寒植物对氮素的利用形态以 NO3−为主[23],高寒草地土壤氮素循环以硝化过程为主,其对系统 N2O 的排放贡献率为 54%[24]。土壤冻融交替的物理撕裂作用,对死亡植物根系起到了破碎作用。土壤微生物类群受控于地上生物量、土壤 C/N 比和 NH+4-N 含量。放牧家畜排泄物的补充,改变了土壤的碳氮比,促进了 N 矿化和硝化基因的增加,反硝化基因的降低,同时由于地上凋落物的移除,土壤的甲烷、碳固持与分解基因被降低,加速了草地生物地球化学循环[25,26],在保障高寒草地养分供求平衡中起到了积极的作用。
海北站站区高寒草地生态系统水分年蒸发散量为 598.0 mm/a,降水量为 472.8 mm/a,且蒸发散中以植被蒸腾为主,占 66.4%,主要受控于大气净辐射[27]。冻土层的“单向”形成,使得深层土壤水分以水汽形态在土表发生表聚,并以冰态储存于土表,季节冻土形成中随水汽迁移的水量贡献为 11% 左右。春季冻土层的“双向”解冻,保障了牧草返青期“春旱”的水分需求,提高了水分的利用率。季节性冻土在高寒草地水分循环中具有特殊作用。
同时,超载放牧干扰下,草地优势植物种群形态上发生低矮化、细绒化和草毡化的演替,以加大啃食难度,增加根系冗余来提高植被的自我恢复能力[28,29]。
尽管温度增加和降水减少都会显著降低植物物种多样性和群落稳定性,温度对群落稳定性的影响,主要体现在降低了组成物种生长的不同步性,降水格局的改变对群落稳定性无显著影响,且群落稳定性的降低与物种多样性并无显著关系[30]。放牧则通过改变牧草的繁殖策略而改变群落结构与多样性[31]。
高寒草甸对于放牧也有较强的适应能力,对相同环境条件下处于不同演替状态的高寒嵩草草甸植物群落生产能力定位监测发现,来自于年际间的变异对植物群落生产能力变异的贡献率为 9.2%,空间异质性对植物群落生产能力变异的贡献率为 7.2%,而放牧干扰强度对群落生产能力变异的贡献率为 83.6%[31]。
未来的全球变暖背景下,高寒草甸的 ANPP(地上净初级生产力)不会受到土壤养分的限制,亦不会引起高寒草甸土壤碳储量改变,适度放牧有利于生态系统的碳周转[25,32]。
研究表明高寒草地生态系统构件通过改善物质能量循环通路,遴选适宜环境条件的植物群落组成结构特征,被动地同环境条件形成完美结合,极大增加了系统的抗干扰和自我恢复能力[33]。重度放牧而非气候变化是引起高寒草地退化的主控因子[34],当前人类赋加于草地的压力远远超过了草地自调控阈值点,这是引起高寒草地退化的主要原因。
3 面向高寒草地生态保护和草牧业发展,通过试验研究和推广应用示范,提高了草地生产力,促进了牧草的高效利用,服务于地方经济建设和人民生活水平提升,对区域畜牧业的发展起到重要支撑作用
20 世纪 70 年代起,海北站就以高寒草地为研究对象,开展了高寒草地承载能力的研究。研究发现高寒草甸草地生产力属中等水平,单个羊单位需求的高寒草甸草地面积为 0.68—1.54 公顷(平均 1.11 公顷);根据中国牧区适用的家畜单位换算系数(每头牛所需草场面积相当于 3.0 个羊单位),每头牛所需高寒草甸草地面积为2.04—4.62 公顷(平均为 3.33 公顷)[35,36]。高寒草甸草场(冬季牧场)适宜载牧率为 3.12 羊单位/公顷(折合 5 亩/羊单位),高寒金露梅灌丛夏季牧场适宜载牧率为 2.67 羊单位/公顷(折合 6 亩/羊单位)。采用划区轮牧和适当延长夏季草场放牧时间,可将该地区草地平均载牧率(9.29 亩/羊单位)提高 35%[37]。
高寒地区自然条件严酷,暖季短暂,牧草生长期仅 100—150 天,产草量较低,而冷季漫长,枯草期长达 7 个月之久,牧草贮存相当匮乏。同时传统的家畜种群年龄结构不合理,畜种比例失调。绵羊种群的牧畜比例一般在 40% 左右,牦牛一般在 35% 左右。羊一般要养到 5 岁以上,母羊 6—7 岁;公牦牛养到 8 岁,母牦牛到 8—9 岁才肯出栏,长寿畜比例很大,严重影响了家畜的繁殖率、出栏率和商品率的提高。在“夏饱秋肥冬瘦”的循环中,浪费了大量的牧草,降低了生产过程中物质和能量的转化效率[38]。3—4 岁母羊每年摄入能量为 9.24×109J,而能量的沉积只有 2.24×108J,平均能量转化效率为 2.4%。该值只占暖季能量利用效率的 15.34%。在冷季,绵羊为维持生命需要消耗 4.31×108J 的体能,相当于摄入总能 2.64×109J。在嵩草草甸地区,每公顷草场可为家畜提供的能量为 2.29×1010J,每只羊在冷季的能量损失,相当于 0.12 hm2草场所提供的能量[37],造成牧草资源的浪费。
针对此现状,海北站进行了高寒牧区家畜合理种群结构及其措施的研究。相应出栏方案是每年秋季出栏全部羯羔羊和老弱羊,并且出栏 3.17% 的成年母羊。在这种方案下,出栏率可提高到 52.79%,平均从每百公斤牧草中收获合 4.9×104J 能量或 3.65 公斤活重的羊产品。从肉用上可取得较大的收益。同时,由于幼羊的出栏,可大大缓解冬季草场压力,有利于季节畜牧业制度的调整。以藏羊为例,家畜的最优结构应为 67.8% 的繁殖母羊,28.36% 的后备母羊,3.84% 的种公羊和后备种公羊[39]。这种家畜结构现已在祁连山区被牧民认可,并得以推广。
由于高寒牧区牧草生产与家畜营养需要的季节不平衡,降低了物质和能量的转化效率,浪费了大量的牧草资源。实行牦牛冬季暖棚育肥是解决草畜矛盾及季节不平衡和保持草地畜牧业可持续发展的主要措施。研究结果表明,冬季在暖棚中采用 35% 燕麦草、20% 菜籽饼、41% 青稞的饲料配方对牦牛进行完全舍饲,其绝对增重、相对增重分别比自由放牧提高 31.44%、26.59%,从而加大牦牛的出栏率和饲草利用率[40]。从经济效益和生态效益综合考虑,牧草利用率为 50% 的情况下,在每年 7—9 月份进行牦牛的育肥,可获得很好的效益。从草地的充分有效利用考虑,在人工草地上放牧育肥犊牦牛,每年 9 月 15 日以前是最佳时期,天然草地放牧育肥牦牛的最佳时期是每年 10 月底以前[41]。在产冬羔的藏系绵羊产区,绵羊生长到 22 月龄屠宰出售可获得较高的经济效益,这时绵羊的胴体重可达 30 斤以上[42]。
受青藏高原独特气候条件的影响。在漫长的冷季,数以万计的家畜死于寒冷,造成严重的经济损失。建造塑料暖棚,改变绵羊越冬生境条件,抗寒保畜具有很高的实用性,是发展牧区畜牧业经济生产的重要途径。棚内冷季 12 月—次年 3 月平均温度(−4.41℃)比同期外界自然环境状况下的温度(−11.76℃)高 7.36℃[43]。温棚的使用使得羔羊的成活率达 95% 以上,同时降低了冬季产热对繁殖母羊的体能消耗。
该项技术已经在青藏高原高寒地区得到了广泛应用与推广,受到牧民和地方政府的高度重视,成为牧区三配套工程的主要内容之一。
通过长期的高原面上高寒草地高寒草甸退化过程与机制的观测,海北站提出了高寒草甸退化状态判别的地方标准[44,45],并以此标准,开展了“家庭制式高寒草地功能提升技术集成与示范”研究。结果发现禾本科牧草为放牧抑制型功能群,莎草科为放牧促进型功能群,对处于两种功能群下的草地应分别采取关键时段延迟放牧、轮牧、减牧或休牧等放牧方法[46]。返青期超载放牧,易导致植物体贮存养分的快速消耗,不利于光合体的拓展和牧草生物量的提高[47];种子成熟期持续超载放牧,易造成种子植物繁殖障碍,导致优势种群演替。植物非生长季放牧对草地的影响较小,适宜的放牧强度应是“吃半留半”。牦牛取食的粗放性、对草地地表践踏强度高,以及对地上植物量移除能力强等因子改变了草地土壤生物结皮所处环境,加速了藻类结皮和地衣结皮的发生、发育、休眠及死亡[48]。对具有较厚草毡表层和致密的土壤生物结皮的草地,应采取草皮的人工划破,以提高种子的着床和萌发率,增加降水入渗,为牧草生长提供适宜生境[49]。草地长期封育,可造成地表凋落物的大量累积,对以种子繁殖的牧草和下繁草的生长造成极大的影响,亦可导致草地的退化,草地的封育年限与其所处演替阶段有关。
养分调控可作为高寒草地植物功能群调控的手段之一。虽然高寒草地植物对 N(氮)和 P(磷)的再利用率分别为 65.2% 和 67.4%,在全球水平上这是最高的,莎草科植物活体 N、P 含量最低,分别为 16.7 mg·g−1和 1.1 mg·g−1,而其再利用率分别为 69.1%和 78.7%[50]。放牧可导致草地土壤表层养分含量的快速下降,随放牧强度的增加,氮素对牧草生长限制变得尤为明显[51,52]。
禾本科牧草为喜氮作物,对氮的需求比较大,氮素的外源补充,可有效提高禾本科牧草的比例和生物量,而嵩草属植物对氮素的补充反应较为迟钝[53]。牧草对养分的需求不仅受到养分含量的制约,同时受到养分配比、养分形态的制约[54,55]。养分的外源添加,不仅改变草地表层的养分状况,还可以引起群落水平物种丰富度的改变、草地地上生产能力的波动,甚至改变表层土壤的酶活性,进而影响到草地生物地球化学自适应调节过程[56]。
过度放牧是导致高寒草地生产功能退化的主导因子,放牧制度的调整是高寒草地功能提升最为有效的措施。海北站的研究表明,放牧制度的选择取决于现实群落优势功能群对牧压的敏感强度和生存策略[31]。对禾草—矮嵩草群落状态的草地恢复措施为采取减牧;矮嵩草群落、矮嵩草+小嵩草镶嵌状态的草地恢复措施为轮牧—休牧;小嵩草群落状态的草地恢复措施为休牧;对杂类草—黑土型退化草地的恢复措施为人工重建[33]。畜群组成的改变(牛改羊)可以作为退化小嵩草草地良化的有效措施[46]。减牧促进了禾本科和豆科植物的生长,禾本科植物相对盖度与生物量显著增加,莎草科和杂类草植物生长受到抑制。禁牧相比自由放牧处理其植被盖度和生物量分别增加 43.4%、39.6%。禁牧 2 年后,草地功能大幅度提升,是非常有效的草地管理措施[47]。
从草地的养分循环、水分利用和种子传播角度来看,天然高寒草地应采取“春季延迟放牧、夏季游牧、秋季轮牧、冬季自由放牧”的放牧制度,对保障草地的可持续利用将起到积极的作用。然而受到家庭牧场草地面积的局限,家庭牧业生产对草地依赖度极高,这种放牧制度难于实施。依据草地所处的演替状态和功能发挥的瓶颈,采取关键生长阶段的草地轮牧,较易被牧户接受。
基于高寒草地适应性管理研究成果,海北站在祁连山区开展了“家庭制式高寒草地功能提升”的技术示范,进行了多期有关高寒草地退化的特征及表观判别、不同退化演替稳态草地的放牧技术、退化草地的恢复与重建等相关专题的牧民培训,培训牧民 300 余人次。并对 6 个牧户草地,实地进行了草地退化状况诊断,针对不同草地退化状态,进行了放牧家畜种类、轮牧方法的现场指导,取得了良好的效果以及牧民的认可(图 4)。
基于退化草地恢复技术成果,海北站在祁连山木里矿区,开展了矿采对草地生态环境、生产力、地表水源传输和渣山植被恢复技术的研究。通过试验、示范,综合恢复效果、投入成本及对环境的二次破坏等因素的考虑,择优选出“整平直播”为矿区受损土地植被恢复的方法,使其恢复成本较政府倡导的“客土覆盖”节约 2.8 万元/亩,在祁连山矿采区生态环境治理起到了引领与支撑作用(图 5)。

图4 家庭制式草地功能提升技术培训

图5 木里煤矿渣山植被恢复技术示范
4 祁连山区是黄河流域重要水源产流地,开展区域生态系统长期监测、生态过程研究、生态系统功能演变评估及其进行功能提升技术集成,超前布局学科重点,为祁连山国家公园体系建设提供科学基础和技术支撑
祁连山是我国西部重要生态安全屏障,是黄河流域重要水源产流地,也是我国生物多样性保护优先区域。放牧作用是祁连山天然草地生态系统主要的土地利用方式;然而,该地区受高原寒冷气候的作用,牧草生长季段、草地生产力低、生态系统脆弱、自我修复能力差。草地特别是位于山前地段的夏季牧场,在长期超载放牧作用下,草地退化严重,不仅造成草地生产功能降低甚至丧失;同时,对山体径流和雪山融水的传输造成极大的影响,造成水源涵养功能的降低。祁连山区还是青海、甘肃两省重要的煤炭矿产地,自 2000 年以来,煤炭资源的开发呈加速趋势,造成区域湿地生态环境的破坏,地表水源传输通道的改变甚至河流的消失,引起国内外的关注。随着祁连山国家公园体系建设工作的展开,高寒草地发生从生产功能向生态功能的转变,以“尊重自然、顺应自然、保护自然”的生态文明理念,保障高寒生态系统的原真性,实现人与自然的和谐发展成为海北站未来服务国家公园体系建设和区域可持续发展学科定位的重点。
海北站在祁连山区已有长达 40 年的定点监测与研究积累,进行了多次青藏高原面上的区域性调查,研究了祁连山高寒草地生态系统的演变及其空间分异特征,提出了基于生态过程的高寒草地适应性分区管理对策。为祁连山国家公园体系建设提供了基础本底数据。同时,针对高寒草地生态系统演变的生物学机制开展生态系统功能评估;建立了高寒矮嵩草草甸系统水平的“放牧干扰实验”控制实验平台、水热改变与养分添加模拟气候变化实验平台和小流域水分收支平台,将开展草地各生物要素对放牧干扰的响应特征及发生的生物学机制研究、草地功能提升及退化生态系统恢复技术集成,为生态建设提供更加准确的数据。
