伊犁河谷苦豆子C、N、P 含量变化及化学计量特征
2018-10-18陈亚宁李卫红朱成刚
崔 东,陈亚宁,李卫红,朱成刚
1 新疆大学资源与环境科学学院/绿洲生态教育部重点实验室,乌鲁木齐 830046 2 中国科学院新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,乌鲁木齐 830011 3 伊犁师范学院生物与地理科学学院/资源与生态研究所,伊宁 835000
植物中碳(C)、氮(N)、磷(P)元素是其所需要的基本营养元素,也是各细胞新陈代谢、蛋白质和遗传物质组成元素。为探索碳、氮、磷等元素的生理生化过程和生物地球化学循环和能量来源起着重要作用,元素的化学计量学的方法是当前生态学研究的热点[1]。宋彦涛、周道玮等[2]相关学者在松嫩草地植物叶片的化学计量学特征的研究上比较充分,近几年来国内外在关于陆地生态系统植物的生态化学计量学的发展较为迅速[3- 4]。如郑艳明等[5]、李红林等[6]、张珂等[7]和陈婵等[8]对陆生植物的根茎叶C、N、P化学计量特征的研究,表明植物在不同生长环境下,能够生长成为适应环境的自身体能机制,从而揭示植物的生存策略。
苦豆子(SophoraalopecuroidesL.)属草本植物[9],主要生于沙质土壤上,具有耐盐碱、抗干旱、耐沙埋等特性,主要分布于我国新疆、宁夏和内蒙古等省区境内。该植物根系较深,地上部分小,地下部分大,形成庞大的根系网。苦豆子强大的根系以及种子蔓延速度极快,使该种群连片生长,扩展性强[10]。在一些严重地区,苦豆子因快速蔓延扩展,可形成苦豆子单优种群。目前,有关苦豆子的研究主要围绕药用价值[11-12],种子形态特征、发芽条件[13- 15]、组织培养[16-17]及种子休眠[18-19]等方面。而关于苦豆子不同器官生态化学计量的研究尚未见报道。伊犁河谷草原苦豆子大面积侵入和连片蔓延生长,已经成为退化草场的优势物种,导致草场质量和经济生产力下降,对新疆伊犁河谷的农田以及草原畜牧业已产生了巨大影响。本研究系统分析和研究苦豆子在不同的季节生长情况下各器官元素的变化规律,有助于了解苦豆子蔓延的生态学机制及对退化草场土壤质量的影响,从而对退化草场的生态恢复提供理论依据。
1 材料与方法
1.1 研究区概况
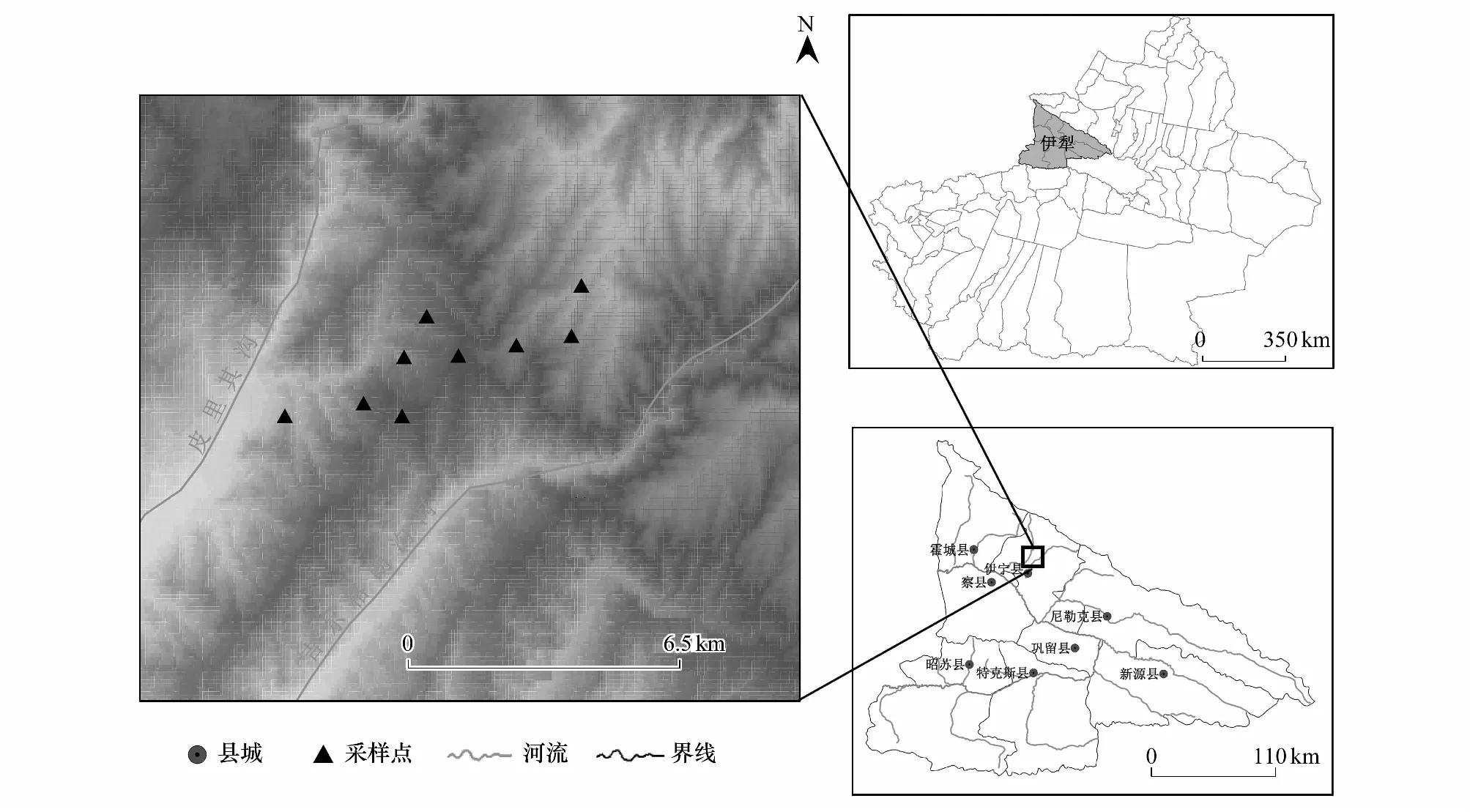
图1 地理位置图和采样点分布Fig.1 Location of the sampling sites
研究样地位于新疆伊犁河谷的托乎拉苏大草原(图1),距伊宁市约40 km(44°01′32″N—44°08′96″N,81°28′40″E—81°35′11″E),属于典型的山地草原地区。这里属于温带大陆性气候半干旱地区,年均气温9—11.1℃,年平均降水250—551.7 mm,年平均蒸发量1621 mm。海拔为900—1700 m。在研究地内,苦豆子多以单优群落的形式分布,群落内伴生植物主要有白羊草(Bothriochloaischaemum)、伊犁蒿(Artemisiatransitiensispoljak)、椭圆叶天芥菜(Heliotropiumellipticum)、毛穗旱麦草(Eremopyrumdistans)、狗牙根(Cynodondactylon)等。
1.2 样品采集及实验分析
在伊犁河谷托乎拉苏草原选择地形、地貌、土壤母质基本一致的9个苦豆子群落作为采样地点,于2016年在苦豆子不同生长期,生长初期,旺季以及生长末期即5月、8月、11月采集样品。在每个样地内随机设定3个1 m×1 m的小样方,在每个样方内剪下苦豆子植物的地上部分,然后将地面枯落物清理干净,挖掘土壤剖面,每个剖面按照每间隔10 cm采集一个植物根系样品(包括根茎和须根),采集深度60 cm,共采集162份植物根系样品以及27个植物地上部分样品。样品在实验室清洗后,烘干称重,粉碎后过筛备用。全碳含量采用重铬酸钾容量法测定、全氮采用凯氏定氮法测定、全磷含量采用钼锑抗比色法测定[20]。
1.3 数据处理
在数据的处理过程中,前期采用Microsoft Excel 2010统计分析,C、N、P采用质量含量,C∶N、C∶P、N∶P采用质量比,后期在SPSS 19.0软件中对不同季节的各不同器官中的碳、氮、磷含量和化学计量比进行单因素方差分析,并采用LSD法进行多重比较。
2 结果分析
2.1 苦豆子碳、氮、磷含量及其化学计量比特征
从表1可知,苦豆子C、N、P含量的极大值分别为699.60、34.98、3.72 mg/g,极小值分别为126.92、3.14、0.47 mg/g,因此苦豆子C、N、P含量的极差较大,分别为572.67、31.84、3.26 mg/g,这与采样时间和不同器官有直接的关系。苦豆子N∶P的最大值为15.37 mg/g,最小值为3.85 mg/g,平均值为8.52 mg/g,平均值小于14,这表明苦豆子的生长主要受到N元素的限制。综合分析苦豆子的变异性可知,变异系数为0.27—0.65之间,属于中等变异。
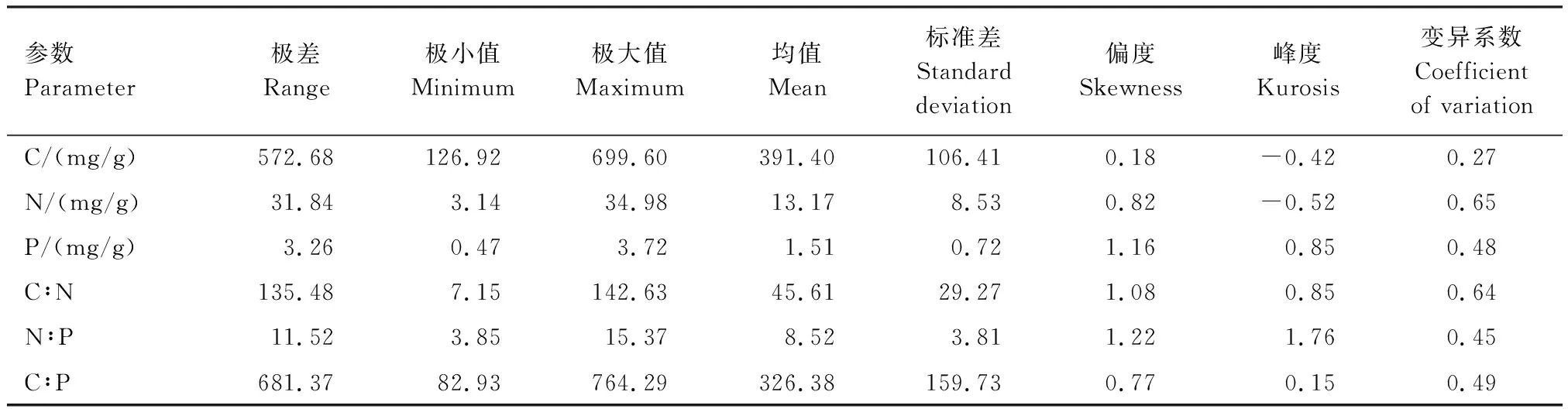
表1 苦豆子C、N、P含量及其化学计量比的统计学参数
从表2可知,苦豆子植物的C含量受生长季节极显著影响(P<0.01)。N含量受器官极显著影响(P<0.01),两因素的交互作用对氮含量的影响也达到极显著水平(P<0.01)。磷含量受生长季节极显著影响(P<0.01),两因素的交互作用对磷含量的影响也达到极显著水平(P<0.01)。
苦豆子C∶N的变异主要受不同器官的影响,其次为季节,两因素的交互作用也达到了极显著水平(P<0.01)。苦豆子植物C∶P主要受到生长季节的限制,其离差平方和为451390.02,为其中的最大值。
2.2 苦豆子不同器官碳、氮、磷含量及其化学计量比特征
在不同时期苦豆子3种器官碳、氮、磷含量具有较大的差异(图2)。各个生长季苦豆子碳、氮、磷含量的变化规律均为叶>茎>根,但随季节变化的动态特征各不相同。苦豆子根、茎、叶中碳的含量在5月的差异性较小,但茎和叶的氮含量要高于根中含量;8月各器官的含量有明显的增加(P<0.05),碳元素的含量达到最高值;根的碳含量表现为8月显著高于5月和11月(P<0.05),茎和叶的碳含量在不同生长时期的差异性显著(P<0.05)。苦豆子叶片中氮含量在5月达到最高值,和8月、11月相比显著性较高(P<0.05)。在5月植物生长发育的初级阶段叶片、根系和茎中的碳、氮值达到峰值,随着生长时间的增加发生递减。
表2苦豆子C、N、P含量及其计量的整体来源变异来源分析
Table2Summaryabouttheeffectofvariationfromdifferentorgans,monthsandbothinteractionsonC,NandPcontentsandtheirratiosofSophoraalopecuroides
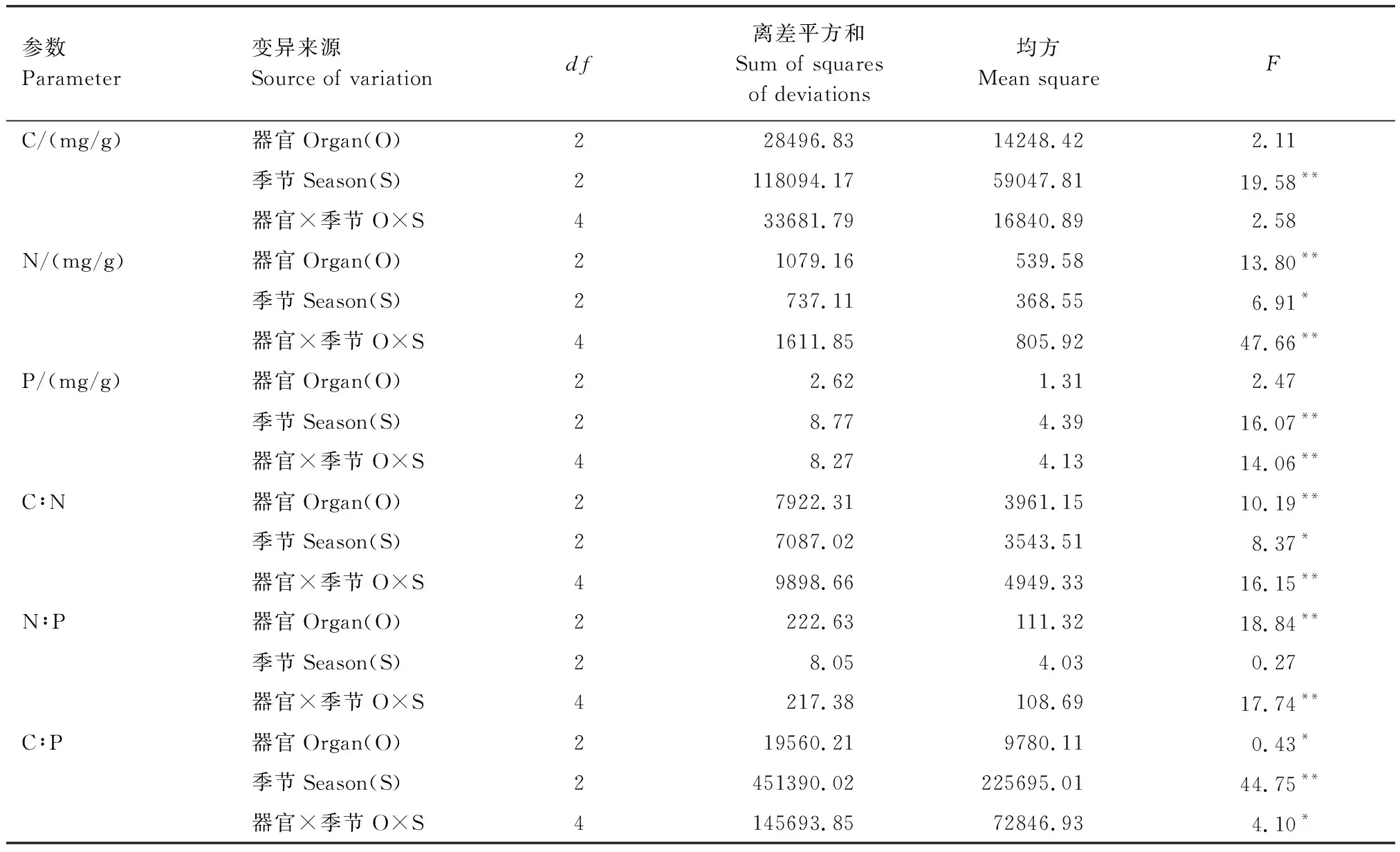
参数Parameter变异来源Source of variationdf离差平方和Sum of squares of deviations均方Mean squareFC/(mg/g)器官Organ(O)228496.8314248.422.11季节Season(S)2118094.1759047.8119.58**器官×季节O×S433681.7916840.892.58N/(mg/g)器官Organ(O)21079.16539.5813.80**季节Season(S)2737.11368.556.91*器官×季节O×S41611.85805.9247.66**P/(mg/g)器官Organ(O)22.621.312.47季节Season(S)28.774.3916.07**器官×季节O×S48.274.1314.06**C∶N器官Organ(O)27922.313961.1510.19**季节Season(S)27087.023543.518.37*器官×季节O×S49898.664949.3316.15**N∶P器官Organ(O)2222.63111.3218.84**季节Season(S)28.054.030.27器官×季节O×S4217.38108.6917.74**C∶P器官Organ(O)219560.219780.110.43*季节Season(S)2451390.02225695.0144.75**器官×季节O×S4145693.8572846.934.10*
**P<0.01,*P<0.05
苦豆子各器官中碳氮磷元素的化学计量比呈现出一定的变化规律(图2)。不同生长时期苦豆子各器官C∶N的变化趋势均为根>茎>叶,5月根C∶N显著高于茎和叶(P<0.05),苦豆子叶片在8月和11月C∶N显著低于根和茎(P<0.05)。5月茎和叶N∶P显著高于根(P<0.05),8月和11月N∶P在根和茎间没有显著差异(P>0.05);根N∶P 在5月和8月间表现出显著性差异(P<0.05),茎N∶P在5月显著低于8月和11月的值(P<0.05)。5月根C∶P显著高于同一时期的茎和叶(P<0.05),8月和11月苦豆子各器官间C∶P差异不显著(P>0.05);不同生长时期的叶片C∶P 差异显著(P<0.05),根和茎C∶P从5月到8月显著增加(P<0.05),之后无显著变化(P>0.05)。

图2 苦豆子各器官碳、氮、磷含量及其化学计量比特征Fig.2 Seasonal dynamics of C,N and P contents and their ratios in different organs of Sophora alopecuroides不同大写字母表示同一器官不同季节间差异显著;不同小写字母表示同一季节不同器官间差异显著( P<0.05)
不同的生长发育阶段对植物各器官的元素含量的积累和分配都有着重要的影响,不同元素的种类对植物各器官的吸收利用过程也不相同[21-22]。分析不同生长季苦豆子各器官C、N、P含量的分配可见(图3),苦豆子植物碳含量在5月各器官的分配比例较为均衡,根茎叶分别为32.08%、34.14%和33.79%;在8月苦豆子碳的含量在各器官的分配比例无明显的波动,与5月相比,根和茎的分配比例降低至31.73%和33.40%,叶中碳含量的分配比例增加到34.87%;11月中根的碳含量有明显的降低为25.26%,茎和叶片中碳含量的分配比例增加到35.54%和39.20%。根和茎中氮含量的分配比随着生长季的增加而逐步减少,根从5月的16.24%降低到生长末期的14.96%;茎从5月的40.65%降低到11月的28.25%;叶片中氮含量的分配比例则不断增加,从5月的43.11%增加到11月的56.80%。根、茎和叶中磷的含量在5月的分配比例分别为21.83%、36.08%和42.08%;在8月的分配比例分别为31.61%、32.88%和35.51%;在11月的分配比例分别为27.44%、27.21%和45.35%。
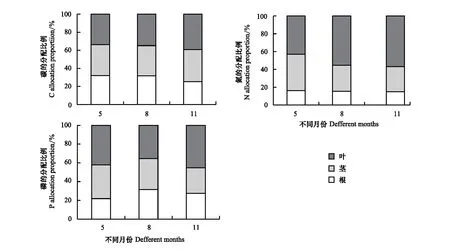
图3 苦豆子根茎叶C、N、P 分配特征Fig.3 C,N and P distribution characteristics of each organ of Sophora alopecuroides
2.3 苦豆子植物根系中的碳、氮、磷随不同深度的变化规律
分析结果显示,苦豆子根系中的全碳、全氮及其全磷含量的变化范围分别为260.80—552.08、3.14—16.34 mg/g和0.37—2.08 mg/g,平均值分别为374.76、7.16 mg/g和1.24 mg/g。根是营养元素的吸收和转运器官,在伊犁河谷苦豆子植物的根系较为发达,在生长季初期根中的氮和磷含量较高,在生长季旺期和生长季末期有明显的降低,两个时期之间的差异较小,因为该时期生长减缓根中的氮磷元素量基本趋于一个稳定值。不同深度的根碳、氮、磷含量变化也有一定的规律性(图4)。随着深度的变化,也显现出递减的趋势。
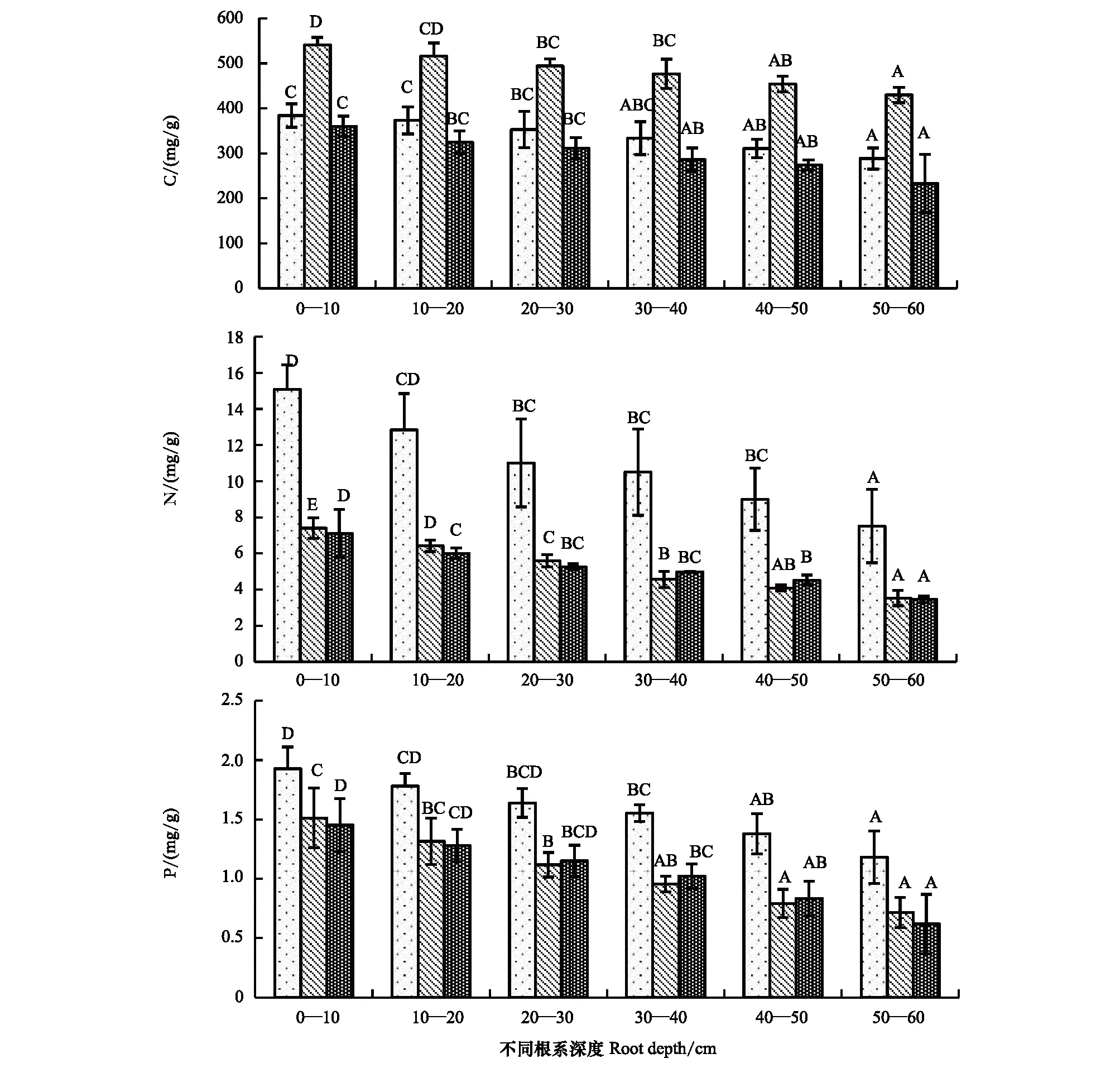
图4 苦豆子不同深度根系碳、氮、磷含量的季节动态Fig.4 Seasonal dynamics of carbon, nitrogen, phosphorusat different depth in root system of Sophora alopecuroides不同大写字母表示同一深度不同季节间差异显著
不同深度的植物根系所能吸收的元素含量不同。观察结果显示,0—10 cm的根系较为发达,C、N、P含量在整个生长季达到峰值,随着深度的增加含量也呈现出递减的趋势,在深度50—60 cm时各含量为最低,根系非常稀疏,土壤中的含量也非常低。在整个生长季内,根系中各元素的含量在生长初期达到最大值,生长末期为最小值。在5月,伊犁河谷内的降水量增加,温度适宜,过渡的放牧使优良牧草减少草场退化,但这给苦豆子植物提供了生长条件和生长空间,使其大肆蔓延,所以根系中的各元素在生长初期达到了最大值。
3 讨论与结论
伊犁河谷的苦豆子C、N、P含量相对较低。C含量变化约为353.95—509.81 mg/g,低于全球492种陆生植物的碳含量[23];P含量为1.61 mg/g,也低于全球P含量水平(1.99 mg/g)。N含量(15.17 mg/g),与全球植物N含量相比也明显较低。苦豆子根、茎和叶C∶N∶P的质量比分别为375∶7∶1、269∶10∶1、226∶11∶1,均都小于全球平均水平。
碳元素是植物生长过程中必不可少的能量元素,一般不会限制植物的生长,氮磷元素是蛋白质和各种遗传物质的重要组成部分,也是植物体内最容易缺少的元素。植物各器官对C、N、P元素的吸收和转化的途径不相同,导致各元素含量在各器官中的分配不同[24]。生长季旺期温度升高,伊犁河谷草原降水量增加,形成优越的自然条件,为苦豆子的大量生长蔓延提供了条件,植物所创造的有机碳含量迅速增加。植物的光合作用增强,叶可以直接获取能源,叶片中含有大量的叶绿素是光和产物的主要器官,在此期间叶片的新陈代谢和生产活动最为迅速,植物体内的生物量增加,碳的含量增多。而在植物的根系中由于细胞膨胀的速度过快,其他器官的氮磷元素减少[25-26]。因此,苦豆子植物茎在转化运输叶片所固定的碳时自身的含量也达到峰值。在生长季的末期植物叶片凋零腐烂回归土壤中,此时,根的吸收能力增强,因此,苦豆子植物碳、氮、磷元素增加达到整个生长季的最大值。
苦豆子碳元素在生长旺季较高,而氮和磷的含量较低,到生长末期碳元素降低而氮和磷元素含量升高。根、茎、叶中的C∶P及C∶N在生长旺期较高,而到了生长末期N、P增加C含量降低,这表明苦豆子采取防御性的生活史策略[27]来增强自身的适应性。苦豆子植物生长初期根中的C∶N、C∶P显著高于茎和叶,这表明在生长初期苦豆子的根部对N和P有较高的利用率,苦豆子主要以根系和种子繁殖为主,在苦豆子的生长初期,苦豆子对N和P较高的利用率可能是苦豆子入侵性强的主要原因之一。在养分贫瘠的环境中,苦豆子靠较高的N、P养分利用策略,在生长初期快速繁殖,占据生长空间,来适应贫瘠的土壤环境。苦豆子生态化学计量特征的季节变化与牛得草等的研究结果相似[28]。伊犁河谷草原苦豆子通过根系和种子进行繁殖蔓延,目前,已经成为托乎拉苏草原退化草场的优势物种,导致草原质量和经济生产力下降。深入研究苦豆子根系碳、氮、磷元素的季节变化,对揭示苦豆子植物在生长过程中地下器官的生长蔓延过程具有重要意义。
