‘金太阳’与‘串枝红’杏正反交后代果实性状的遗传倾向研究
2018-10-18武晓红景晨娟陈雪峰赵习平袁立勇张宪成唐焕英李立颖
武晓红,景晨娟,陈雪峰,赵习平*,袁立勇,梁 爽,张宪成,唐焕英,李立颖
(1.河北省农林科学院 石家庄果树研究所,河北 石家庄 050061;2.河北省玉田县林业局,河北 玉田 064100;3.河北省巨鹿县农林局,河北 巨鹿 055250;4.河北省顺平县林业局,河北 顺平 072250)
杏为蔷薇科(Rosaceae)杏属(Armeniaca)植物,原产于我国,其种质资源极为丰富[1]。杏成熟早,外观美,风味佳,是我国人民喜爱的时令水果,现已成为世界性水果之一[2]。近30年来,我国十分重视杏种质资源的收集和研究工作,通过有性杂交选育出了一批杏新品种[3-6]。但中国的杏品种多数自交不亲和,遗传上高度杂合,制约了我国杏育种和生产的发展[7]。遗传研究是育种的重要基础,深入了解遗传变异规律,有助于正确选配亲本,制定合理的育种目标,获得理想的杂种后代[8]。迄今为止,关于杏果实性状的遗传研究已取得了一些进展[9-11],但多以自然杂交或是单一杂交组合后代群体为研究对象,且有些果实性状的遗传规律还没有明确;有关同时对正反交后代果实性状的遗传倾向进行分析的研究还未见报道。
本试验以欧洲早熟杏品种‘金太阳’、河北极晚熟地方杏良种‘串枝红’以及它们的正反交后代为材料,探讨了杏果实发育期、平均单果重、果形指数、果实盖色、果实硬度、可溶性固形物含量、可食率等果实性状的遗传规律,旨在为杏杂交育种的亲本选配及后代果实性状表型的预测提供依据。
1 材料与方法
1.1 试验材料
以在河北省农林科学院石家庄果树研究所种质资源圃栽培的20年生‘串枝红’与‘金太阳’为亲本,以‘金太阳’ב串枝红’为正交,‘串枝红’ב金太阳’为反交。于2010年杂交,2011年播种,2012年定植,2014年开始结果。随机选取正交后代114株、反交后代81株实生树,于2016年开展调查。亲本的株行距为3 m×5 m;杂交后代采用宽窄行定植,株行距为0.8 m×1 m×2 m。果园土质为沙壤土;在树体生长期间采用相同的施肥、灌溉、喷药、疏果、修剪等管理措施。
1.2 试验方法
待果实成熟后每株随机选取10个果,采回至实验室进行测定。测定指标:杂交后代及亲本的果实发育期、单果重、果实横径、果实纵径、果实侧径、果皮盖色、果实硬度、可溶性固形物含量、鲜核重、核横径、核纵径、核侧径等;测定按照《杏种质资源描述规范和数据标准》[12]执行。
单果质量和单核质量用PTY-B5000 1/100天平测定。果实纵、横、侧径和核纵、横、侧径用游标卡尺测量。果实硬度测定用LANDTEK FHT-05数显果实硬度计,测定部位为果实胴部,去皮面积约1 cm2。可溶性固形物含量采用Atago PAL-1数显糖度计进行测定,将10个果实分别捣烂,取汁液进行测量。
相关计算公式如下:果形指数=果实纵径/0.5×(果实横径+果实侧径);核形指数=核纵径/核横径);可食率=(单果重-鲜核重)/单果重;变异系数CV(%)=(杂种后代标准差/杂种后代平均值)×100;遗传传递力Ta(%)=杂种后代平均值/中亲值×100;优势率H(%)=[(杂种后代平均值-中亲值)/双亲平均值]×100。
2 结果与分析
2.1 金太阳与串枝红正反交后代果实发育期的遗传变异
金太阳为早熟品种,果实发育期为69 d;串枝红为极晚熟品种,果实发育期100 d。两个亲本进行正、反交,后代果实发育期的平均值小于中亲值(表1)。在正交后代中有87.72%单株的果实发育期介于双亲之间,在反交后代中有93.83%单株的果实发育期介于双亲之间。正、反交后代的遗传传递力分别为93.31%和96.43%。可见杏正反交后代的果实发育期呈趋中的遗传倾向,果实发育期有较高的遗传力,该性状的表现受环境因素的影响较小。正、反交后代果实发育期的变异系数分别为10.11%、11.71%,说明正反交后代果实发育期的变异程度较大。正、反交后代的超高亲比率分别为1.75%和7.47%,低于低亲比率分别为10.53%和3.70%。以金太阳做母本时,更易于将其短果实发育期性状遗传给后代。

表1 金太阳与串枝红正反交后代果实发育期的遗传变异
2.2 金太阳与串枝红正反交后代果实大小的遗传变异
试验结果(表2)表明:正反交后代的平均单果重均小于双亲的平均值,正交后代甚至低于低亲;正交后代低于低亲的比率达90.35%,反交后代为55.56%,遗传传递力分别为75.39%和92.86%,可见杏正反交后代单果重有趋小的遗传倾向;正反交后代单果重的平均遗传传递力为84.13%,遗传力较小,表现易受环境因素的影响;正、反交后代单果重的变异系数分别为24.33%和13.59%,可见正反交后代单果重的变异程度较大。
2.3 金太阳与串枝红正反交后代果形指数的遗传变异
从表3可以看出:金太阳与串枝红正、反交后代果形指数的平均值均大于中亲值,遗传传递力分别达到101.79%和100.28%,变异系数分别为5.36%和6.75%,可见果形指数的遗传力较强,变异程度较小;正、反交后代果形指数超高亲比率较高,分别为49.12%和48.15%,可见金太阳与串枝红正反交后代果形指数均有变大的遗传倾向。
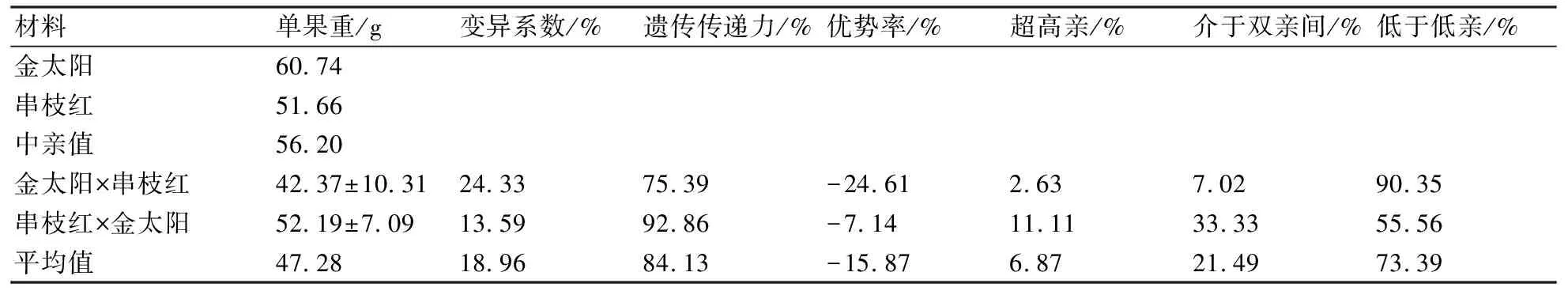
表2 金太阳与串枝红正反交后代果实大小的遗传变异
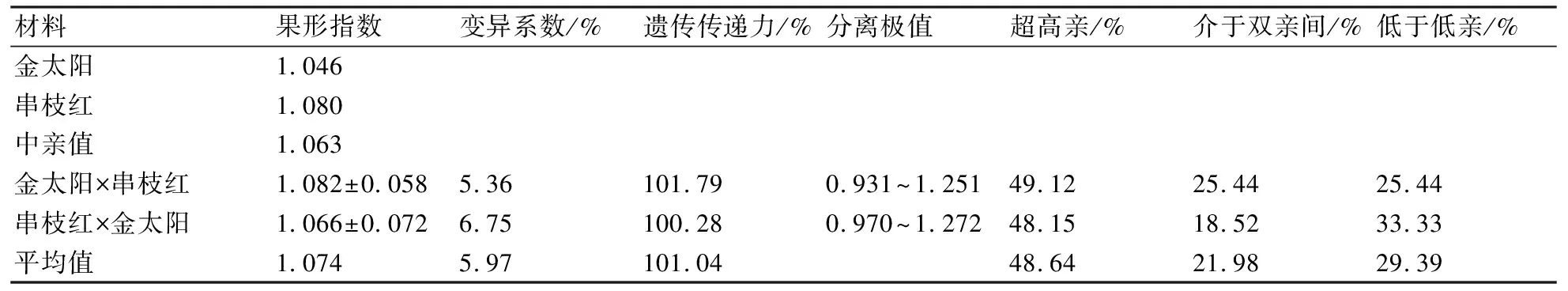
表3 金太阳与串枝红正反交后代果形指数的遗传变异
2.4 金太阳与串枝红正反交后代果皮盖色的遗传变异
金太阳底色橙黄色,无彩色;串枝红底色橙黄,着2/4~3/4紫红色。双亲正反交后,果实盖色出现了紫红、红、粉红以及无彩色的分离(图1)。在正交后代中,无彩色的杂种单株为83株,占72.81%;着粉红色的为18株,占15.80%;着红色的为9株,占7.89%;着紫红色的为4株,占3.51%。在反交后代中,无彩色的为42株,占51.85%;着粉红色的为30株,占37.04%;着红色的为6株,占比7.41%;着紫红色的为3株,占比3.70%。正反交后代的果皮盖色均以无彩色为主,紫红色最少。可见,金太阳的无彩色更易遗传给后代,但是以串枝红做母本时,杂交后代果皮着彩色的比率较高。
2.5 金太阳与串枝红正反交后代果实去皮硬度的遗传变异
从表4可以看出:金太阳与串枝红正反交后代的果实去皮硬度均大于中亲值,遗传传递力分别为112.73%和100.91%,说明果实硬度的遗传力较强;正、反交后代果实硬度介于双亲间的比率分别为55.26%和59.26%,表现为趋中变异;但在正、反交后代中也分别有27.20%和18.52%的超高亲杂种单株,这为今后的杂交育种提供了素材;正、反交后代果实去皮硬度的变异系数分别为48.39%和47.45%,变异程度较大;正、反交后代果实去皮硬度广泛分离,分离极值分别为6.31和4.29。
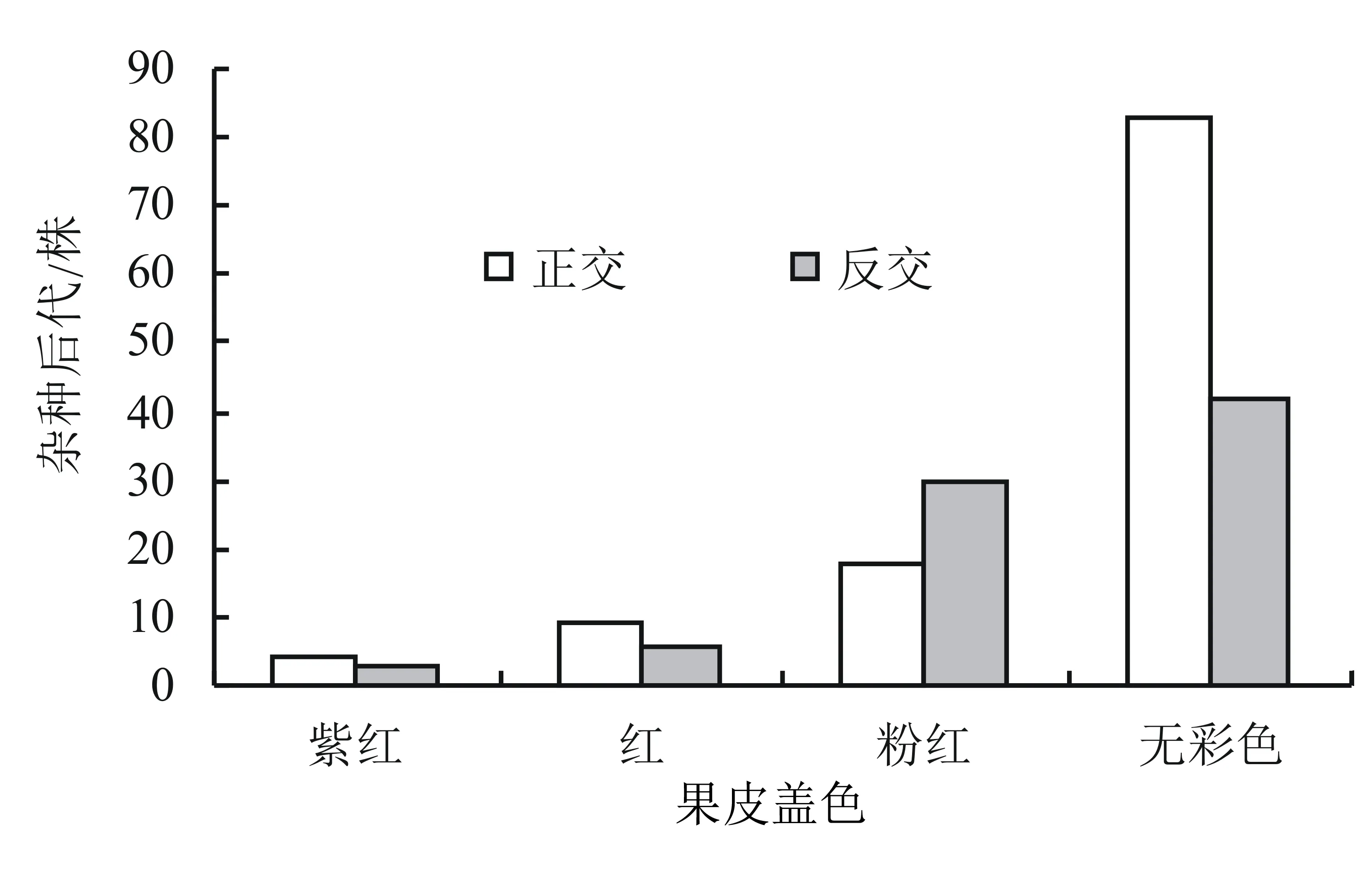
图1 杂种后代果皮盖色的分布
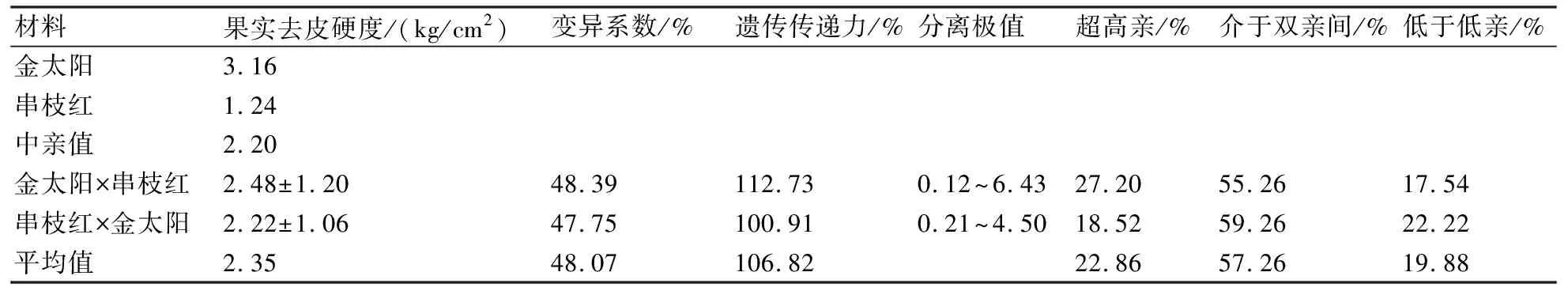
材料果实去皮硬度/(kg/cm2)变异系数/%遗传传递力/%分离极值超高亲/%介于双亲间/%低于低亲/%金太阳3.16串枝红1.24中亲值2.20金太阳×串枝红2.48±1.2048.39112.730.12~6.4327.2055.2617.54串枝红×金太阳2.22±1.0647.75100.910.21~4.5018.5259.2622.22平均值2.3548.07106.8222.8657.2619.88
2.6 金太阳与串枝红正反交后代果实可溶性固形物含量的遗传变异
从表5可以看出:金太阳与串枝红正、反交后代的果实可溶性固形物含量平均值均大于中亲值,遗传传递力分别达到108.04%和101.30%,变异系数分别为21.27%和21.89%,可见,可溶性固形物含量的遗传力较强,变异程度较大;正、反交后代果实可溶性固形物含量介于双亲间的比率分别为64.04%和74.07%,呈趋中变异;后代分离程度较高,分离极值分别为15.0和13.3。

表5 金太阳与串枝红正反交后代果实可溶性固形物含量的遗传变异
2.7 金太阳与串枝红正反交后代果实可食率的遗传变异
从表6可以看出:正、反交后代果实的可食率平均值均低于双亲平均值,正交后代的可食率平均值甚至低于低亲值;正、反交后代中低于低亲比率分别为76.32%和40.74%,遗传传递力分别为98.56%和99.79%,变异系数分别为1.72%和0.96%。可见,正反交后代的果实可食率有减小的遗传倾向,且组合遗传传递力较高,变异系数较小。果实可食率有较高的遗传力,表明该性状的表现受环境因素的影响较小。
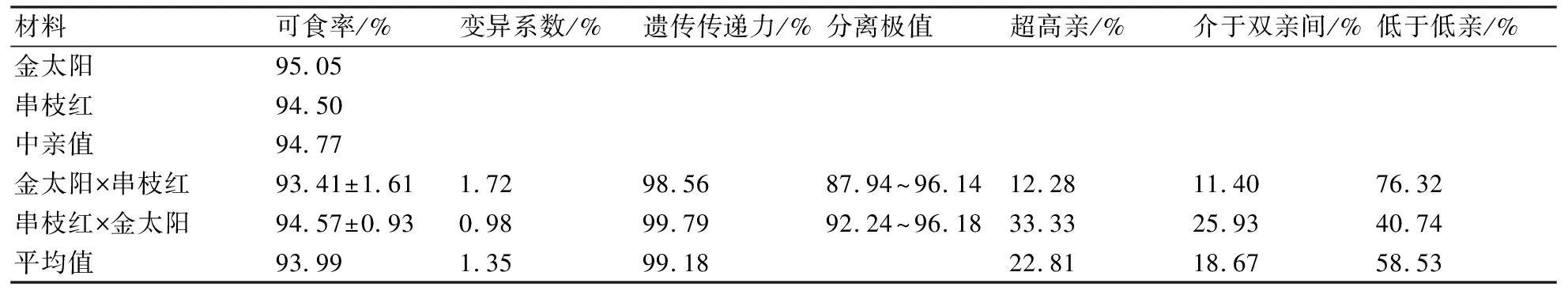
表6 金太阳与串枝红正反交后代果实可食率的遗传变异
2.8 金太阳与串枝红正反交后代果实核形指数的遗传变异
从表7可以看出:正、反交后代果实核形指数的平均值均小于中亲值,介于双亲间的比率分别为79.82%和77.78%,遗传传递力分别为94.12%和94.85%。可见杏正反交后代果实的核形指数呈趋中的遗传倾向,该性状的遗传力较强,受环境因素的影响较小。正反交后代果实核形指数的变异系数分别为7.03%和7.75%,变异程度中等。

表7 金太阳与串枝红正反交后代果实核形指数的遗传变异
2.9 金太阳与串枝红正反交后代果核厚度的遗传变异
从表8可看出:正、反交后代果实的果核厚度平均值均低于双亲平均值,遗传传递力分别为96.85%和94.49%;正交后代中果核厚度介于双亲之间的比率为44.74%,反交后代中低于低亲的比率为66.67%,可见串枝红小的果核厚度性状遗传给后代的能力较强;正、反交后代果核厚度的变异系数分别为65.85%和77.50%,,变异程度较大。
3 讨论
在杏有性繁殖的后代中,亲本性状的非加性效应解体,会出现复杂的分离,后代会出现综合双亲优良性状或产生新性状、超亲性状等以供选择。本研究结果表明,金太阳×串枝红正反交后代果实发育期呈趋中变异,这与前人的研究结果[13-14]一致。当以果实发育期较短的金太阳做母本时,低于低亲比率达到10.53%,说明当以短果实发育期的亲本做母本时更易将短果实发育期性状遗传给后代。果实大小是由多基因控制的数量性状[8]。本研究发现:杏正反交后代的单果质量均呈普遍变小的趋势,低于低亲的平均比率为73.39%,这与赵习平等的研究结果[9]一致;正反交后代单果质量均出现了超亲单株,但比率较低。正反交后代果实的可食率遗传趋势与单果质量的遗传趋势相似,呈现趋小的遗传倾向,可见单果重变小的趋势大于单核重变小的趋势。因此,要想选育出大果类型或可食率高的类型,还需要以大果类型或可食率高的类型作为亲本。果形是一个比较复杂的性状,杂种后代有广泛的分离[15]。在本研究中,金太阳与串枝红正反交后代果形指数的超高亲率分别为49.12%和48.15%,果形指数变大的遗传趋势明显;其正、反交后代果形指数的变异系数分别为5.36%和6.75%,分离程度都不高,双亲的这一性状的非加性效应较弱。

表8 金太阳与串枝红正反交后代果核厚度的遗传变异
一般认为,果实表色是由少数基因控制的显性或不完全显性性状[8]。本研究结果表明,金太阳与串枝红正交后代中有72.81%为无彩色杂种单株,反交后代中有51.85%为无彩色杂种单株,在有果实盖色的单株中,从粉红色到紫红色的单株比率逐渐降低。杏果实盖色的无彩色对彩色为显性。刘振中[16]、盛炳成等[17]研究认为,苹果果皮表色由主效基因控制,不仅由一个或两个简单的显性基因支配,还由一个或多个受生长条件影响的辅助基因控制。花色苷是杏果皮呈现红色的原因[18]。花色苷颜色的表现及其稳定性受到温度、光照、糖等诸多因素的影响[19],可见,在基因型和环境条件的共同作用下,杏果实表皮的红色性状以不同程度显示。
果实硬度与贮运性成正比,选育果实硬度大的杏品种是当前的育种目标之一。在本试验中,杏正反交后代果实去皮硬度的平均值均大于中亲值,非加性效应在该性状的遗传中起主要作用。正、反交后代果实去皮硬度介于双亲间的比率分别为55.26%和59.26%,趋中变异明显;但是高于高亲的比率分别为27.20%和18.52%,且遗传传递力较大,说明果实去皮硬度受环境的影响较小,受遗传的影响较大,可通过杂交育种选育果实硬度较高的类型。
一般认为,果实可溶性固形物含量呈明显的数量性状遗传,并表现出超亲优势[20]。本研究结果表明,金太阳与串枝红正、反交后代中果实可溶性固形物含量介于双亲间的比率分别为64.04%和74.07%,呈趋中变异,且平均值均大于中亲值,表明金太阳与串枝红正反交后代的可溶性固形物含量有增高的趋势;其遗传传递力分别达到108.04%和101.30%,变异系数分别为21.27%和21.89%。可见,果实可溶性固形物含量的遗传力较强,变异程度较大,通过本杂交组合可以选育出高可溶性固形物含量的后代。
本试验结果表明,金太阳与串枝红正反交后代果实核形指数呈趋中的遗传倾向,果核厚度呈趋小的遗传倾向,可见该性状的遗传力较强,受环境因素的影响较小。
本研究还显示,正、反交后代果实的果核厚度平均值均低于双亲平均值,遗传传递力分别为96.85%和94.49;正交后代中果核厚度介于双亲之间的比率为44.74%,反交后代中低于低亲的比率为66.67%,可见串枝红小的果核厚度性状遗传给后代的能力较强。
