亚热带侵蚀红壤区植被恢复过*程中土壤团聚体化学计量特征
2018-10-17区晓琳陈志彪陈志强陈海滨任天婧
区晓琳 陈志彪† 陈志强 陈海滨 任天婧
(1 湿润亚热带山地生态国家重点实验室培育基地,福州 350007)
(2 福建师范大学地理科学学院,福州 350007)
土壤团聚体是土壤结构的基本单元,通常由不同尺度矿物颗粒和胶体物质共同参与发生凝聚胶结作用结合形成大小不一的多孔介质结构体。土壤团聚体是土壤碳、氮、磷等养分的存储与转化场所以及土壤微生物的生存环境,对调控植物生长、降低水力侵蚀、提高土壤肥力水平等有重要作用[1]。由于不同粒径团聚体形成方式和胶结物质不同,它们对有机碳、氮和磷的吸附形态以及稳定性存在差异,导致对养分固定及转化能力的不同[2-3],影响着土壤养分地球化学循环过程。生态化学计量学作为对生态系统养分循环研究的重要工具,是分析多重元素,尤其是结构性元素(C)与限制性元素(N、P)之间的平衡关系以及生态系统交互影响的理论依据[4-5],对植物、凋落物、土壤及微生物等碳氮养分循环、养分限制、碳固定以及森林演替等起指示性作用[6]。土壤作为一个复杂的生态系统,土壤中碳、氮、磷元素是植物生长重要的养分来源,主要储存在不同粒径团聚体内,植被状况对团聚体养分固持与循环的影响不容忽视。目前,关于不同植被状况下团聚体研究以团聚体形成机制、稳定性、养分尤其是碳氮固存较多[7-8],对植被恢复过程中土壤团聚体中碳、氮、磷生态化学计量特征的研究相对较少。
由于地理环境以及植被破坏的影响,福建省长汀县水土流失区成为我国中亚热带花岗岩红壤生态系统脆弱区的典型代表。境内陡坡丘陵山地集中分布,土壤抗蚀性极差,且保水保肥能力低下,植被的自然恢复较为困难,且稀疏的植被覆盖加剧土壤侵蚀发生,面蚀、沟蚀和崩岗等侵蚀地貌广泛分布[9]。植被恢复是实现生态修复的重要对策,由于恶劣的生态环境,自然植被恢复过程存在较大的困难,采用人工植被恢复措施是加快生态恢复过程的重要手段。自20世纪80年代以来,该地区相关部门相继实施一系列以乔-灌-草恢复植被措施,经过30多年恢复,该区生态恢复成效显著。植被恢复必然导致侵蚀红壤中碳氮磷元素的输入/输出产生深刻的影响,通过生态化学计量特征可表征该地区养分循环特征。目前,已有学者针对植被恢复过程土壤碳矿化[10]以及生态化学计量特征[11]等进行比较系统的研究,土壤团聚体方面亦有研究[12],但对植被恢复对侵蚀退化地土壤团聚体碳、氮、磷及其化学计量特征的认识尚浅。因此,本文通过化学计量学理论,以福建省长汀县典型侵蚀退化红壤区中不同植被恢复年限侵蚀坡地为研究对象,探讨植被恢复过程中不同粒径团聚体有机碳、全氮和全磷含量变化及其生态化学计量特征,分析植被恢复与侵蚀红壤团聚体养分化学计量特征的关系,对于揭示红壤侵蚀区植被恢复过程中土壤团聚体中碳、氮、磷的平衡关系以及土壤团聚体碳氮“汇”功能变化有重要指示性意义,为定量评价红壤区生态恢复效果提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于闽西南长汀县河田镇朱溪小流域(116°23′30″—116°30′30″E,25°38′15″—25°42′55″N),属中亚热带季风湿润气候,年均气温18.5 ℃,年均降雨量1 710 mm,年均蒸发量1 403 mm,年均无霜期260 d,大于等于10 ℃积温4 100~4 650 ℃。土壤类型以燕山运动晚期中粗晶花岗岩风化发育形成的红壤、侵蚀性红壤为主,粗砂粒含量较高,抗蚀性极差;由于长期的土壤侵蚀和人为干扰,常绿阔叶林为主的地带性植被遭到严重破坏,几乎被马尾松(Pinus massoniana Lamb.)次生林替代。自20世纪80年代以来,研究区采取种植生态(林)草植被恢复措施,在保留“老头松”的基础上,补植木荷(Schima superba Gardn.et Champ.)、枫香(Liquidambar formosana Hance.)、胡枝子(Lespedeza bicolor Turcz.)等乡土树种,辅植宽叶雀稗(Paspalum wettsteinii Hackel.)、香根草(Vetoveria zizanioides Linn.)及百喜草(Paspalum natatu Flugge.)等草本植物。
在全面野外勘查的基础上,依据典型性和代表性为原则,考虑成土母质和立地条件等综合因素,选取未治理地(0 a)作为对照,恢复年限分别为5 a、10 a、15 a、30 a以及80 a(该样地为次生林,树龄约为70~90 a,平均为80 a),共计6处典型样地。除恢复80 a样地外,其余5个样地在植被恢复前,土壤母质均为花岗岩,土壤A层剥蚀殆尽,B层厚度仅为5~10 cm,地形条件、侵蚀状况和人为扰动均相似。未治理地(恢复0 a)虽经过几十年时间自然恢复,地表土依然裸露,植被稀少,浅沟发育,坡面侵蚀严重;恢复5 a样地坡顶部分基岩裸露,凋落物较少,植被以马尾松、胡枝子、枫香和芒萁为主;恢复10 a和15 a样地地表由芒萁覆盖,凋落物厚度分别为0~3 cm、0~5 cm,植物群落以马尾松、黄檀(Dalbergia hupeana Hance.)、芒萁(Dicranopteris dichotoma(Thunb.)Bernh.)、金茅(Eulalia speciosa)、五节芒(Miscanthus floridulus(Labill.)Warb.)为主;恢复30 a样地林下植被稀少,凋落物厚度3~8 cm,植物群落以杉木、马尾松、胡枝子和芒萁为主;恢复80 a样地林下小型灌木较多,凋落物厚度3~10 cm,植物群落以马尾松、枫香、荷花玉兰(Magnolia grandiflora Linn.)等为主。各样地基本理化性质见表1。

表1 不同恢复年限样地基本状况Table 1 Basic situations of six plots different in restoration history
1.2 样品采集与测定
2015年8月上旬进行野外调查与采样,为降低天气因素对土壤状况扰动,采样前15天,天气均以晴天和多云为主,气温在23~36℃之间。在每个样地内设置3个20 m×20 m的标准样方,相邻样方间距约10 m,近等高线平行布设样方。样方内按“S”形设置5个采样点,并分0~20 cm和20~40 cm两个土层采集原状土样,同一土层的5点土样等量混合装入1个硬质塑料盒内,共采集原状土样36份。原状土样在室内自然风干后,沿自然结构轻轻掰成直径约1 cm的小土块,除去植物残体、砾石等杂物,采用四分法,取四分之一土样,用于土壤理化性质的测定,剩余的原状土样采用沙诺维夫干筛法分离出>5 mm、2~5 mm、1~2 mm、0.5~1 mm、0.25~0.5 mm、<0.25 mm 共6级土壤团聚体组成后,分别研磨过0.149 mm筛,用于有机碳、全氮和全磷的测定。
土壤容重采用环刀法测定;土壤含水量采用铝盒烘干法测定;土壤pH按水土2.5︰1采用STARTER 300便携式酸度计测定。土壤团聚体有机碳和全氮采用碳氮元素分析仪(Elemantar vario MAX, 德国)测定;全磷用硫酸-高氯酸消煮定容及过滤,提取待测液后采用连续流动分析仪(Skalar san++, 荷兰)测定。
1.3 数据处理与分析
土壤碳氮比(C︰N)、碳磷比(C︰P)和氮磷比(N︰P)均采用质量比。数据经Excel 2003初步整理后,统计分析采用SPSS 19.0进行,采用单因素方差分析(One-Way ANOVA)确定不同恢复年限各粒径土壤团聚体化学计量变化特征,显著性检验采用Duncan 新复极差法(显著性水平设为α=0.05),利用Pearson 相关系数评价土壤团聚体养分及其化学计量特征之间相关程度,图表制作通过Origin 9.0完成,图表数据均以平均值±标准差表示。
2 结 果
2.1 植被恢复过程土壤团聚体养分含量变化
侵蚀退化生态系统经植被恢复后,土壤团聚体中的养分贮量出现不同程度的增加。由图1可知,土壤团聚体有机碳、全氮和全磷含量变化范围分别为2.06~27.71 g·kg-1、0.50~2.12 g·kg-1和0.034~0.189 g·kg-1。各粒级团聚体有机碳、全氮和全磷含量总体表现为随植被恢复年限增加呈显著增加趋势;随团聚体粒径减小,有机碳、全氮和全磷含量总体上表现为增长趋势。
0~20 cm土层中,在植被恢复过程中,各粒级团聚体有机碳含量表现为随着恢复年限的增加呈显著升高的趋势,到恢复80 a时,各粒径团聚体有机碳含量增加了4.93倍~6.27倍。有机碳含量随团聚体粒径减小表现为显著增加特点(除恢复0 a 各粒径有机碳变化不明显外,其他随着年限的增加,各粒径间的有机碳含量越来越显著),其中<0.25 mm粒径有机碳含量是>5 mm粒径的1.66倍~3.31倍。团聚体全氮含量亦表现为随植被恢复年限增加呈增加趋势,其中恢复80 a是恢复0 a的2.44倍~3.42倍。除恢复0 a和5 a 全氮含量随粒径减小而增加不显著外,其他年限团聚体全氮含量均表现出随着粒径减小而显著增加特点(P<0.05)。全磷含量在植被恢复过程呈升高趋势,变化大小则表现为80 a >30 a >15 a >10 a >0 a >5 a。除了恢复前15 a的全磷含量随粒级减小变化不明显外(P>0.05),其他年限各粒级团聚体全磷含量则随粒级减小略有增加趋势,其中< 0.25 mm粒径团聚体全磷含量显著高于>1 mm粒径(P<0.05)。
20~40 cm土层亦受到植被恢复的影响,土壤团聚体有机碳、全氮含量分配特征与表层0~20 cm相似,但含量大小和增加幅度明显低于表层,0~20 cm土层的团聚体全磷含量亦略高于20~40 cm土层,三者均表现出养分的表层富集特征。恢复前15 a各粒级团聚体有机碳含量随年限增加而无显著增加(P>0.05),恢复30 a和80 a则表现显著增加趋势。各粒级有机碳含量随粒径减小而显著增加(恢复0 a除外)。团聚体中的全氮含量表现为在植被恢复前、中期间(恢复30 a之前)无明显差异(P>0.05),恢复80年则呈显著升高趋势。植被恢复过程中,各粒级团聚体全磷含量呈增加趋势,变化的大小顺序为80 a >10 a >30 a >15 a >0 a >5 a。随粒径减小其全磷含量略有升高,但粒径间无显著差异(P>0.05)。
2.2 植被恢复过程土壤团聚体生态化学计量特征
由图2可知,植被恢复过程中,各粒径团聚体C︰N变化范围为3.06~13.05,随植被恢复年限增加,各粒级C︰N总体呈升高趋势,随粒径减小总体上呈增加特点。0~20 cm土层中,团聚体C︰N变化范围为4.49~13.05,随恢复年限增加,各粒径C︰N呈显著升高趋势,到恢复80 a时,C︰N升高了1.84倍~2.32倍。随团聚体粒径减小,C︰N呈显著升高趋势(P<0.05),其中<0.25 mm粒径的C:N是>5 mm粒径的1.24倍~1.86倍。相对于0~20 cm土层,20~40 cm土层的团聚体C︰N值略低,在3.06~11.72之间。在植被恢复过程中,团聚体C︰N表现为升高→降低→升高的趋势,其中恢复80 a的C︰N值最高,恢复15 a的C︰N值最低,恢复80 a的C︰N是恢复15 a的1.65倍~2.25倍。团聚体C︰N随粒径减小总体呈升高趋势,除恢复0 a各粒径团聚体C︰N无显著差异(P<0.05),其他恢复年限中<0.25 mm粒径的C︰N均显著高于>5 mm粒径(P<0.05)。
由图3可知,植被恢复过程中,各粒径团聚体C︰P变化范围为21.4~185.6,表层0~20 cm各粒径团聚体C︰P明显高于20~40 cm土层。随恢复年限增加,各粒级的C︰P总体上表现呈升高→降低→升高趋势,随粒径减小,C︰P总体上呈增加趋势。0~20 cm土层,C︰P的变化范围为51.16~185.6,各恢复年限的土壤团聚体C︰P表现为80 a>5 a>10 a>15 a>30 a>0 a,恢复80 a是恢复0 a的1.68倍~2.78倍。在不同恢复年限各粒径中,除恢复30 a团聚体C︰P随粒径减小无显著差异(P>0.05),其他恢复年限的团聚体C︰P随粒径减小则呈显著增加趋势(P<0.05)。20~40 cm土层的总体上各粒径团聚体C︰P低于0~20 cm,其变化范围为21.40~158.90,团聚体C︰P表现为80 a >5 a >0 a>30 a>10 a>15 a,其中恢复80 a是15 a的2.46 倍~2.94倍。除恢复0 a和15 a中团聚体C︰P在各粒径中无显著差异外(P>0.05),其余年限的团聚体C︰P则随粒径减小总体上呈显著增加趋势(P<0.05)。
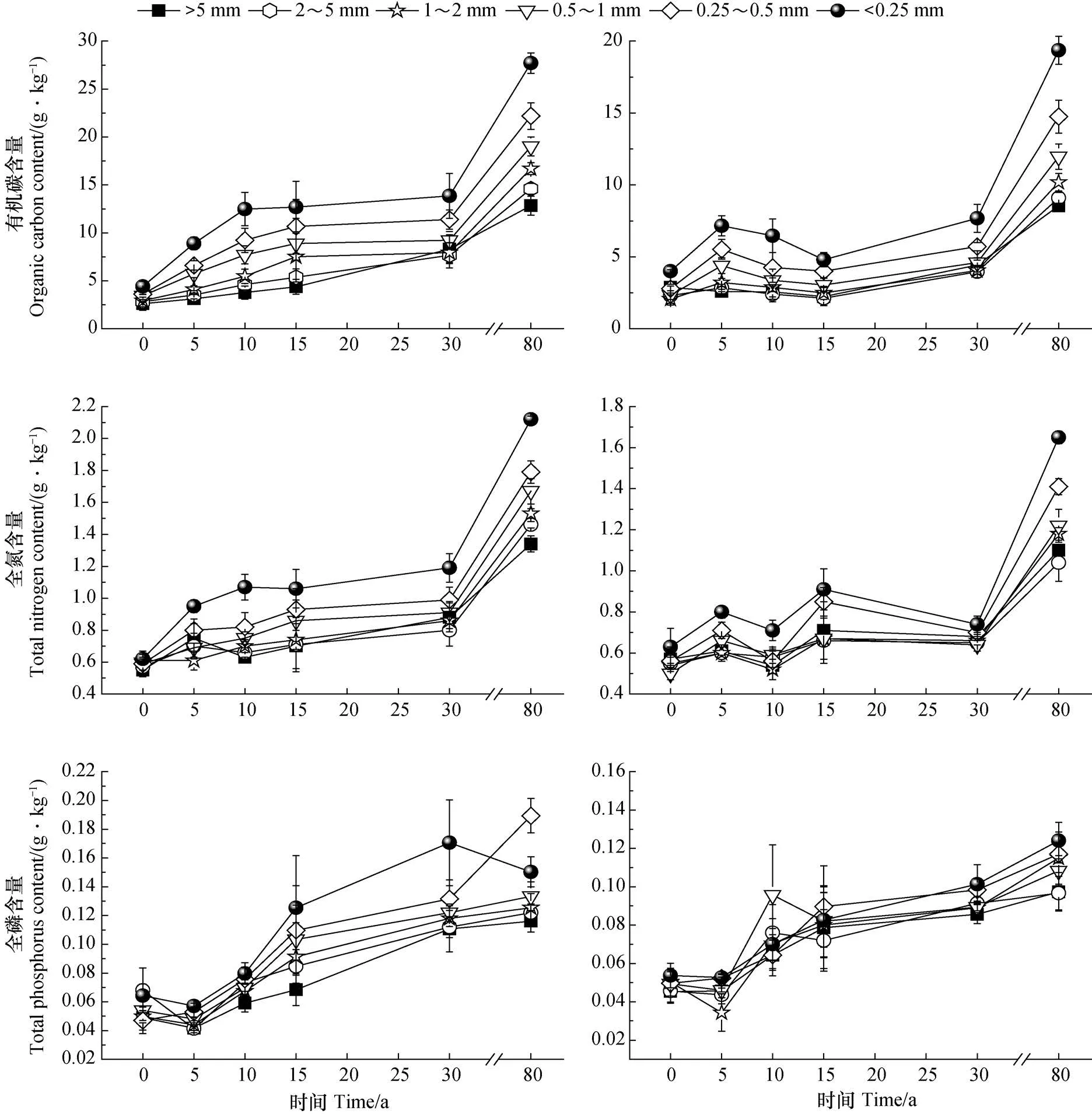
图1 植被恢复过程土壤团聚体有机碳、全氮和全磷含量分布特征Fig. 1 Distribution characteristics of organic carbon, total nitrogen and total phosphorus contents in soil aggregates relative to vegetation restoration process
由图4可知,植被恢复过程中,各粒级团聚体N:P变化范围为5.62~18.20,两土层间团聚体N︰P变化不大。随着恢复年限增加,团聚体N︰P呈增加→减小→增加趋势,恢复5 a时,团聚体N︰P最高,恢复30 a时N︰P最低。随粒径减小,团聚体N︰P总体上呈先减小后增加的趋势,但各粒径间无显著差异(P>0.05)。0~20 cm土层内,N︰P变化范围为7.18~18.20,恢复5 a中各粒径团聚体N︰P显著高于其他年限(P<0.05),是恢复30 a的1.91倍~2.45倍。20~40 cm土层的N︰P值和分布特征及大小与0~20 cm土层相似,N︰P的变化范围在5.62~15.13,恢复5 a是30 a的1.67倍~2.09倍。两个土层中,不同恢复年限各粒级随粒径减小N︰P无显著变化(P>0.05)。
2.3 植被恢复过程土壤团聚体养分及化学计量特征的关系
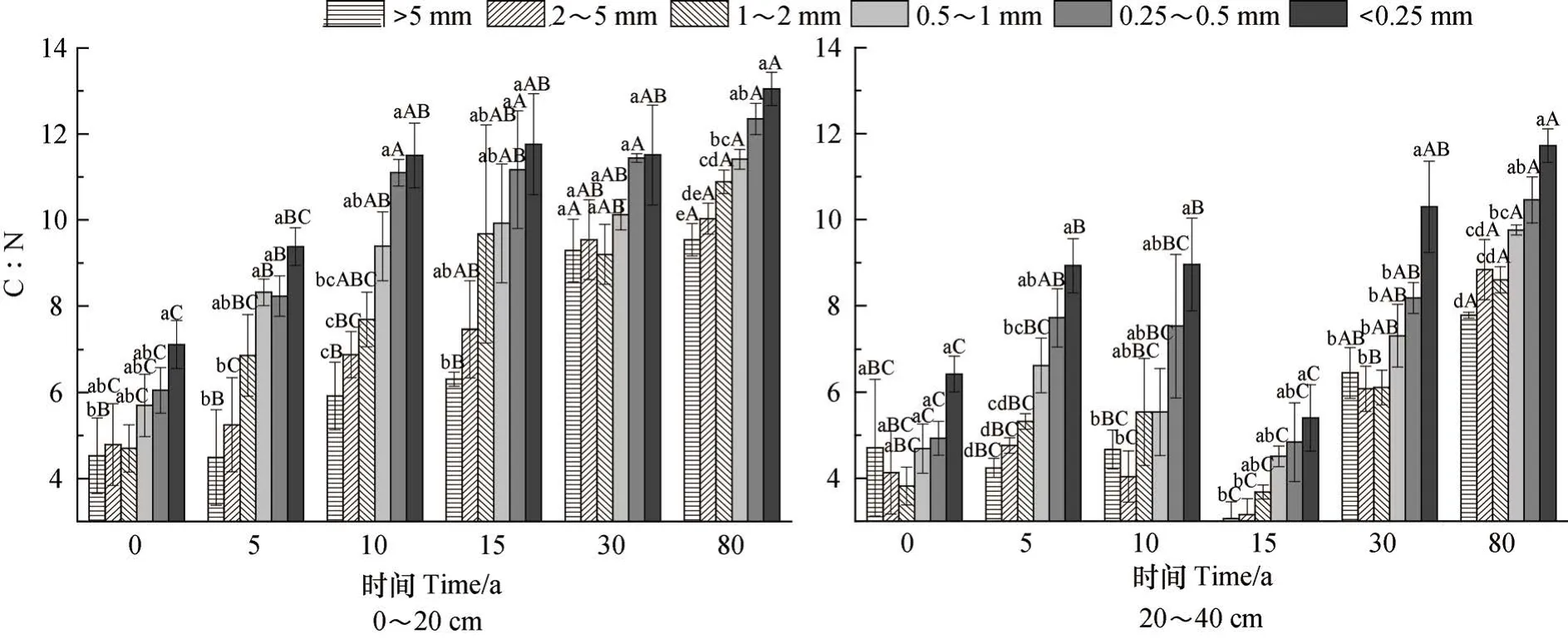
图2 植被恢复过程土壤团聚体C:N化学计量特征Fig. 2 Stoichiometric characteristics of C:N in soil aggregates relative to vegetation restoration process
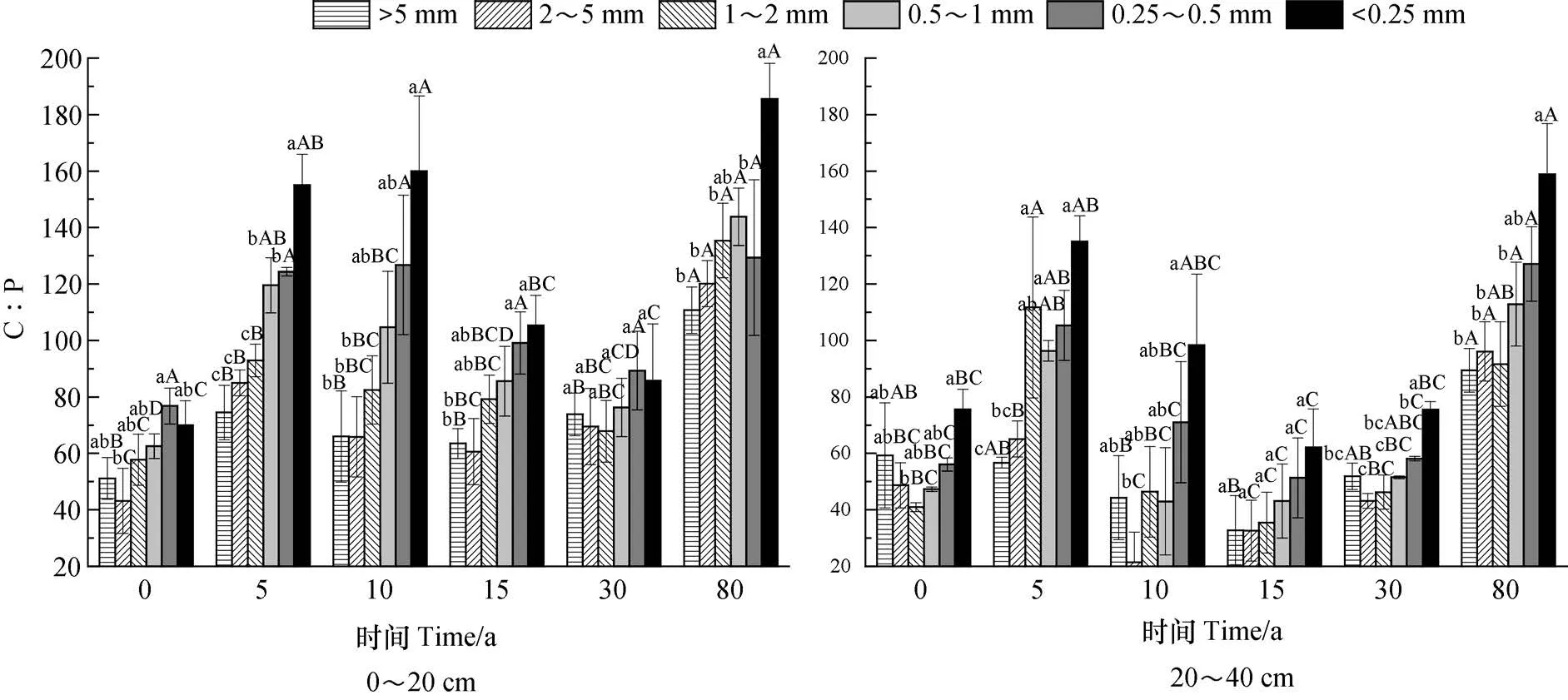
图3 植被恢复过程土壤团聚体C︰P化学计量特征Fig. 3 Stoichiometric characteristics of C:P in soil aggregates relative to vegetation restoration process
为进一步了解土壤团聚体养分及化学计量的相互影响,对植被恢复过程中侵蚀红壤团聚体养分及化学计量进行相关性分析(表2)。结果表明,土壤与团聚体有机碳、全氮和全磷存在不同相关程度,土壤C、N、P元素分别与团聚体中C、N、P呈极显著正相关(P<0.01),相关范围分别为0.83~0.93、0.84~0.92、0.67~0.75。随粒径的减小,土壤与各粒径团聚体C、N元素的相关程度总体上呈升高趋势,土壤与各粒径团聚体P元素则随粒径减小总体上呈降低趋势。
不同粒径团聚体有机碳、全氮和全磷三者之间含量均存在极显著正相关关系(P<0.01),各粒径团聚体C与N元素的相关程度达0.9以上,且随粒径减小呈升高趋势。各粒径团聚体C与P的相关系数在0.65~0.76,各粒径团聚体N与P的相关系数在0.60~0.76。团聚体中C和N元素与团聚体C:N均为极显著正相关(P<0.01),且随粒径减小相关性呈增加趋势。各粒径团聚体C与C:N的相关系数明显高于N与C:N的相关系数。不同粒径团聚体C与C︰P呈极显著正相关(P<0.01),且随粒径减小相关性呈降低趋势。不同粒径团聚体P与C︰P相关系数偏低(0.13~0.29),且无明显相关性。不同粒径N与N︰P相关系数在0.11~0.29,且无明显相关性,不同粒径团聚体P与N︰P呈极显著的负相关(粒径为<0.25 mm为显著负相关),各粒径间的相关性变化较小(<0.25 mm除外)。
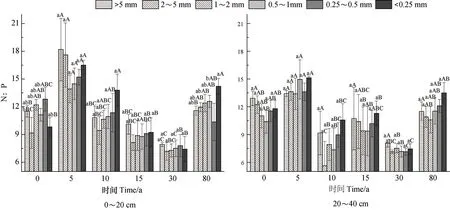
图4 植被恢复过程土壤团聚体N:P化学计量特征Fig. 4 Stoichiometric characteristics of N:P in soil aggregates relative to vegetation restoration process

表2 土壤团聚体养分及化学计量相关性Table 2 Correlations of nutrients and stoichiometry in soil aggregates
3 讨 论
3.1 不同恢复年限对土壤团聚体养分的影响
土壤团粒结构作为土壤有机质的保持场所,土壤中约90%的有机碳储存于土壤团聚体[13],土壤团聚体对固碳和改善土壤肥力十分重要。本研究中,各粒级团聚体有机碳、全氮和全磷含量随植被恢复年限增加而升高,这与以往的研究结果一致[12,14-15]。0~20 cm土层团聚体有机碳和全氮含量明显高于20~40 cm土层,且随恢复年限增加,土层间差异越明显,出现养分聚表特征。这主要由于植被凋落物是土壤有机碳和全氮的主要输入途径[16],植被恢复有效地增加侵蚀红壤表土层动植物残体、根系分泌物以及微生物等有机质,改善团聚体结构,还降低了土壤侵蚀强度,减少土壤养分流失。在植被恢复过程中,除了恢复80 a表土层(0~20 cm)团聚体有机碳和全氮含量(18.84 g·kg-1、1. 65 g·kg-1)与福建省[17]表土层(0~20 cm)红壤的有机碳、全氮含量平均水平(22.1 g·kg-1、1.49 g·kg-1)相近,团聚体全磷含量(0.124 g·kg-1)略低亚热带未侵蚀山地[17](次生林)土壤全磷含量(0.15 g·kg-1)外,其他恢复年限的团聚体养分含量均低于恢复80 a,表明亚热带退化地经过80 a的植被恢复后土壤团聚体有机碳、全氮含量基本可恢复到亚热带未侵蚀土壤的平均水平。土壤中磷元素是主要来源于地质岩石风化,南方红壤其磷素含量主要与成土过程、风化特征以及土壤侵蚀相关因素相关,同时也受动植物残体分解的影响[18]。低纬度地区强烈的风化淋溶作用引起土壤中全磷含量普遍偏低,加之,本研究区曾遭受严重的土壤侵蚀,磷素早已淋溶殆尽,全磷含量极低[18]。研究发现[19],地表凋落物的分解提高土壤有机磷素的累积,从而提高土壤磷的总储量。因此,随着植被恢复年限增加,地表枯枝落叶层增厚,通过微生物的分解矿化土壤团聚体中全磷含量亦随着植被恢复时间的延长而增加。但其含量不高,总体仍低于亚热带未侵蚀红壤地区的平均水平,甚至远远低于全国全磷含量(0.65 g·kg-1)[20]。
关于植被恢复对土壤团聚体养分分配特征存在不同的研究结论。王景燕等[21]对川西退耕5 a坡地研究发现,团聚体有机碳和全氮含量随粒径的减小呈“V”形变化趋势;孙娇等[22]对黄土高原不同林龄刺槐土壤研究发现,团聚体有机碳、全氮和全磷含量的粒级分布呈两边低中间高的趋势。本研究中,土壤团聚体有机碳、全氮、全磷含量总体上随着粒径的减小呈增加趋势,这与前人相关研究结果一致[14,23-24]。研究地随植被恢复年限增加,粒径越小,有机碳、全氮和全磷含量提高越明显,主要由于较小粒级团聚体比表面积大,吸附有机质能力强,不易被土壤微生物分解[15],表明较小粒径团聚体对土壤养分的富集能力更强。关于土壤团聚体养分分配结论存在较大差异可能与不同成土母质化学组成差异、土壤类型、植被条件、侵蚀状况及人为活动影响等因素相关。此外,从相关性分析可看出,土壤C、N、P元素分别与团聚体C、N和P元素呈极显著相关,表明团聚体中C、N、P元素对土壤养分状况均具有良好的指示作用,且粒径越小,对土壤C、N的指示性更强,这可能由于土壤有机质是储存和调控养分有效性的库,并受到各粒径团聚结构不同程度的保护[25]。
3.2 不同植被恢复年限对土壤团聚体化学计量特征的影响
土壤化学计量特征是土壤有机碳与氮素、磷素之间的平衡耦合机制,土壤碳氮磷元素的输入/输出之间的平衡耦合及其有效性决定生态系统碳循环和碳固定效率[4]。受气候、地貌、成土母岩、植被条件等自然环境调控和人类活动的干扰,土壤中养分含量及循环速率不同,造成C︰N︰P有较大的空间变异[4],本文重点讨论植被状况与团聚体C︰N︰P的关系。表土层(0~20 cm)各粒级团聚体C︰N、C︰P、N︰P与全国土壤C︰N、C︰P、N︰P(均值分别为11.9、61、5.2)以及热带和亚热带的C︰N、C︰P、N︰P(均值分别为12.1、78和6.4)相比[20],除恢复80 a表土层团聚体C︰N与全国土壤C︰N水平接近外,其余恢复年限的团聚体C︰N的总体水平均低于全国和热带与亚热带的平均水平。较低的C:N意味着土壤中有机质分解矿化速率快,养分循环周期短,这可能与退化生态系统植被表现出较强环境适应性有关[8]。同时,团聚体C︰N低表明碳氮“汇”能力较差,这不利于土壤团聚体碳氮养分的固存。随着植被恢复年限增加,土壤各粒级团聚体C︰N呈显著升高特点,这与李玮等[23]研究不同植茶年限各粒级团聚体C︰N无显著差异的结果存在差异,且与以往的研究中认为C、N元素作为结构性成分,两者在累积与消耗过程中存在比较稳定的比值[4,26]的结论有所不同,可能由于研究区曾为极度侵蚀退化地,土壤中的碳、氮循环遭受破坏,土壤生态系统养分循环处于不稳定状况,虽然进行了植被恢复措施以提高土壤养分,但有机碳、氮输入/输出仍然未达到平衡状况,可能还需要更长时间修复退化生态系统以达到碳氮比值较稳定状态。此外,通过相关性分析还发现,团聚体C、N元素与团聚体C:N呈极显著正相关,而C与N 亦极显著正相关,即团聚体C和N元素对环境的响应几乎同步,但可能由于植被恢复过程中团聚体C元素增加速率大于N元素的增加速率,团聚体C对团聚体C:N的影响更为强烈。总之,土壤团聚体C:N随植被恢复呈增加趋势,提高土壤碳氮的固存量,增加土壤团聚体碳氮“汇”功能,对改善土壤肥力乃至对全球气候变化有重要意义。
土壤中C︰P通常被认为是土壤磷素矿化能力的标志或是从环境中吸收固持磷素潜力的一种标志[27]。土壤C︰P比值低表明微生物在矿化土壤有机质中释放磷的潜力较大,C︰P比值高则表明微生物对土壤有效磷有同化趋势[6],具有较强的固磷潜力。在生态恢复过程中团聚体C︰P表现为升高→降低→升高变化特征,未治理前表层土壤团聚体的C︰P最低(60.23),恢复30 a时表层团聚体C︰P与亚热带相近,其他恢复年限均远远高于亚热带平均水平。造成这种差异可能由于研究区未治理前遭受强度侵蚀,土壤养分流失殆尽,团聚体中的有机碳和全磷含量均极低,较低团聚体C︰P水平却有利于微生物在分解有机质过程中释放团聚体中的有效磷,加之侵蚀地植物生长缓慢,对磷素的需求量相对较低,团聚体中的有效磷素基本上能满足植物生长需求。恢复中前期(5 a、10 a),可能由于植被迅速生长,需要大量的磷元素,在分解有机质过程中出现微生物与植物竞争土壤无机磷的现象[27],磷素被固定在团聚体中,导致团聚体C︰P值呈升高趋势,而植物生长缺乏磷素,不利生长。恢复80 a,团聚体C︰P为最大值(131.3),表明植被恢复后期土壤中微生物释放磷素严重不足,不能满足植物生长需求,这与李玮等[23]研究植茶后期土壤团聚体磷素有效性下降的结果相似。
土壤中的氮、磷元素是植物生长所需的必需矿质元素以及是生态系统中最常见的限制性元素,土壤中N︰P可作为土壤养分限制类型的有效预测指标[4]。随植被恢复年限的增加,土壤团聚体中N︰P变化表现为增加—减少—增加,但其值均高于全国和热带与亚热带水平。在恢复0 a到5 a时,土壤团聚体N︰P迅速升高,这可能与植被恢复初期,对补种植物施加氮肥的生态林草措施,促使土壤中氮素含量升高有关。从恢复5 a到30 a,土壤团聚体N:P值虽有所下降,但土壤团聚体N︰P整体水平仍然较高,并高出亚热带土壤N︰P的平均水平[20],而在以往的研究表明亚热带地区总体受P元素的限制[28],且在植被恢复过程中,侵蚀红壤团聚体N︰P值一直居高不下,土壤团聚体明显受到P元素的限制。此外,从团聚体N、P与团聚体N︰P的相关性分析中亦可发现,土壤团聚体P元素与团聚体N︰P极显著负相关关系,与土壤团聚体N元素则无显著相关,表明土壤团聚体N︰P主要受P元素的影响。因此,在植被恢复过程中,由于团聚体中P元素增长缓慢,难以满足植物生长需求,团聚体中的P元素成为南方侵蚀红壤植被恢复过程主要的限制性元素。
不同粒径团聚体化学计量特征存在明显差异,即不同粒径团聚体对土壤养分的供应能力亦不同。植被恢复过程中,土壤团聚体C︰N随粒径减小而升高,主要由于凋落物等有机质进入土壤后,有机质首先进入较大粒级团聚体中,由于较大粒级团聚体中新有机碳结构简单,稳定性较差,有机碳容易微生物被分解矿化[26],这对土壤退化生态系统恢复过程中,较大粒级土壤团聚体有机碳的矿化能短期内有效地为植物生长提供养分供植被吸收利用,这对侵蚀红壤地的生态恢复有积极的意义。而较小粒级团聚体有机碳由于受物理保护作用[13],容易被固定。在植被恢复过程中小粒径团聚体的C︰N增加速率远高于大粒径团聚体,表明随植被恢复年限增加土壤团聚体中有机碳存储能力增强,尤其是小粒径团聚体碳储能力远大于大粒径团聚体[23]。土壤团聚体C︰P随粒径减小而升高,表明较大粒径团聚体中微生物分解有机质释放磷的潜力高于较小粒径团聚体,即较大粒级团聚体更容易矿化释放磷素,而较小粒径团聚体利于磷素储存。由于各粒径N︰P值均较高,明显受到磷元素的限制,不同恢复年限各粒级土壤团聚体N︰P随粒径减小表现为稍有减少后增加趋势,但粒径间无显著差异,表明较大粒级土壤团聚体和微团聚体均受到磷元素的限制较大,但各粒径间磷元素的限制程度不明显。
4 结 论
随着生态恢复年限的增加,红壤侵蚀区各粒径土壤团聚体中有机碳、全氮和全磷含量以及C:N呈显著升高趋势,而团聚体C:P、N:P变化不稳定,表现为升高→降低→升高趋势;随土壤团聚体粒径减小,未治理地各粒径养分无显著差异,其它恢复年限团聚体有机碳、全氮和全磷含量均随着粒径的减小总体上表现为升高趋势,团聚体C︰N和C︰P总体上呈显著升高趋势;表土层(0~20 cm)的团聚体有机碳、全氮含量以及C︰N、C︰P明显高于底土层(20~40 cm),表土层团聚体全磷含量略高于底土层,且随恢复年限增加两个土层间团聚体养分差异越明显,团聚N:P在两个土层间无明显差异。从团聚体养分与化学计量的相关性来看,团聚体碳、氮、磷元素对土壤养分具有良好指示性;团聚体碳、氮、磷元素具有同质效应;团聚体C、N元素影响着C︰N,但C元素影响力更大;团聚体P元素对C︰P的影响更明显,团聚体N︰P主要受P元素的影响。以上结果表明,植被恢复显著提高了侵蚀红壤团聚体中有机碳、全氮和全磷含量,团聚体中碳氮“汇”功能增强,较小粒径团聚体对碳、氮、磷元素的固持能力大于较大粒级团聚体,团聚体中P元素成为退化生态系统恢复的限制性元素。
