模拟增温对玉米叶片生理特性及根区土壤特性的影响
2018-10-16张其国周旭丹王忠东
李 苇,张其国,周旭丹,王忠东
(1.吉林农业大学园艺学院,吉林长春130118;2.白城职业技术学院,吉林白城137000)
100年来,全球气候变化正改变着生态系统平衡,全球平均温度已上升了0.6℃,带来了持续性的负面影响,因此成为21世纪影响全球农业生产的重要环境因素之一[1-3]。大量野外模拟增温的研究表明,20世纪全球范围内的温室效应正在继续和蔓延,根据2013年的IPCC评估报告,到21世纪末,预计全球平均气温将升高1.8~4.0℃[4]。这种高温可能会对农作物的新陈代谢和土壤代谢过程产生复杂的影响[5]。温度控制着土壤生态系统中大量的微生物代谢和反应速率,并起到调节生态系统能量和养分的作用,在一定程度上改变了农作物的产量,从而间接地改变土壤生态系统过程[6-7]。因此,在全球气候变化背景下,农作物对气候条件所表现出的适应和生存方式对其生长、生态及土壤特性产生了深远影响。
玉米具有产量高、营养丰富、生态适应性广等优良特点,同时对于调节气候和改善农业生产具有极其重要的意义[8-10]。在全球气候变化背景下,研究玉米的生长特性及土壤特性对全球气候变化的响应,不仅可以加深对其生态适应、生理特性、生产潜能的系统性认识,而且为玉米的高产优质栽培提供理论依据。因此,针对全球气候变化背景下玉米生长特性不够明晰的状况,试图在前人研究的基础上,利用开顶式生长室(open-topchamber,简称OTC)进行小尺度生态系统模拟控制试验,并连续3年同步监测模拟增温对玉米生长特性及土壤特性的影响,从生长生理特性、土壤特性等方面揭示玉米对全球气候变化的响应机制,为实现全球气候变化背景下玉米高产优质栽培提供试验数据支撑。
1 材料与方法
1.1 试验材料
供试玉米品种为沈糯3号,研究区位于吉林农业大学野外试验田,分别于2014—2016年在玉米整个生长季进行增温处理。该区多年平均气温5.6℃,多年平均降水量750 mm。供试土壤质地为壤质黏土,耕层土壤黏粒含量25.84%,土壤pH 值 6.76,有机碳含量 20.41 g/kg,全氮含量 1.89 g/kg。
1.2 试验设计
采用随机裂区设计,设置对照(CK)和模拟增温(OTC)2个试验处理,每个处理各6个重复,共12个试验小区。模拟增温小区面积为5.0 m×5.0 m,由约2.0 cm厚氟化玻璃制成,高4.0 m,顶部开口后略收紧(以减少气体交换),通过红外辐射加热管进行模拟增温处理,每个开顶式生长室设置1根红外辐射加热管(功率220万W),24 h不间断增温,每个增温装置正面处的温度差值≤0.2℃,增温效果均一,增温在5℃左右;对照无红外辐射加热管。于2014年,在沈糯3号的整个生长季进行增温处理。在OTC内与对照CK小区设置Fluke 2638A全能型温度数据采集器,在每个生长季选取晴朗无风的1天,每隔1 h测定OTC和CK地上和地下10 cm的地温。将直径15 cm、高12 cm的土壤呼吸底座(PVC)埋入各个小区边缘,定期除去底座内的杂草,为减少对环境的影响,在观测土壤呼吸速率前1个月将PVC测量圈埋入土壤中,以为保证足够的时间平衡其生态环境。
1.3 测定项目及方法
2014—2016年8月底,分别在OTC和CK小区内,按照0.5 m×0.5 m收割地上生物量,并采集足够多的成熟叶片带回实验室,65℃烘干至恒质量(精确到0.001 g)。同时采用四分法采集0~15 cm原状土壤(≥2 kg),速封袋保存带回实验室,去除石块、动植物残体和大的颗粒型物质,过2 mm筛,4℃ 保存备用。
1.3.1 玉米农艺性状 不同时期的每个小区选取5~10株长势一致的玉米,测量株高、茎粗,扫面仪测定植株叶面积指数,然后烘干叶片测定干质量,计算比叶重;在成熟期收获后测定百粒质量、穗粒数和穗粒质量[11]。
(1)不同时期在每个小区选取5~10株长势一致的玉米,卷尺和游标卡尺测量株高、茎粗(精确到0.01 cm),用扫描仪测定植株叶面积指数,并在成熟期收割晒干后称干质量。(2)将单株玉米根系挖取出来,每珠地上与地下用塑料袋分开,带回实验室,40目筛网流水冲洗,65℃烘干后测定干质量。RIS=植株地下部质量/植株地上部质量。(3)不同时期采集的玉米叶片烘干粉碎,采用混合液浸提法测定叶绿素含量(CCM-200叶绿素仪);采用蒽酮法测定碳水化合物含量,采用考马斯亮兰法测定蛋白质含量,碳水化合物和蛋白质的含量以占玉米叶片干质量的百分比表示;采用氮蓝四唑法测定超氧化物歧化酶(SOD)活性;采用愈创木酚显色法测定过氧化物酶(POD)活性;采用紫外分光光度法测定过氧化氢酶(CAT)活性;采用硫代巴比妥酸法测定丙二醛含量[12]。
1.3.3 土壤酶活性 在玉米成熟期,采用四分法采集每个小区玉米根区0~15 cm原状土壤(≥2 kg),用速封袋保存带回实验室,去除石块、动植物残体和大的颗粒形物质,过2 mm筛,4℃冰箱保存备用测定土壤酶活性。其中,土壤蔗糖酶活性采用3,5-二硝基水杨酸比色法测定;土壤脲酶活性采用苯酚钠比色法测定;土壤酸性磷酸酶活性采用磷酸苯二钠法测定;土壤转化酶活性采用分光光度法测定[13]。
1.3.4 土壤呼吸速率 分别在不同时期选取晴朗无风的一天,从07:00开始到16:00结束,采用动态密闭气室红外CO2分析仪(IRGA)法(美国Li-6400便携式气体分析系统和Li-6400土壤呼吸室)每隔3 h测定1次土壤呼吸速率。
1.4 数据统计分析
数据采用Excel 2003和SPSS 18进行统计分析,采用单因素方差分析(One-way ANOVA)和t检验比较各处理间的差异显著性。
2 结果与分析
2.1 模拟增温对玉米生长发育的影响
2.1.1 生长特性 由表1可知,模拟增温显著影响玉米生长期的特性,玉米株高、茎粗、叶面积指数、比叶重和单株生物量在不同时期均表现为OTC>CK,而R/S表现为OTC<CK;随着生长期的进行,OTC和CK处理下玉米株高、茎粗、叶面积指数、比叶重和单株生物量均呈逐渐增加的趋势,而OTC和CK处理下玉米R/S呈逐渐降低的趋势;其中OTC与CK之间的差异随生长期的进行而逐渐拉大,在成熟期二者均达到极显著差异水平(p<0.01),除了单株生物量,OTC与CK处理下其他生长指标在分化期差异均显著(p<0.05),也即增温处理促进了玉米的生长发育,并促成了地下生物量的累积,其中在成熟期的促进作用表现得最为明显。

表1 模拟增温对玉米植株生长特性的影响
2.1.2 生理生化特性 由表2可知,模拟增温显著提高了玉米叶片叶绿素a、b和碳水化合物、蛋白质含量,在不同时期均表现为OTC>CK。随着生长期的推进,OTC和CK处理下玉米叶片叶绿素a、b和碳水化合物、蛋白质含量均呈逐渐增加的趋势,其中OTC与CK之间的差异逐渐拉大,在成熟期二者均达到极显著差异水平(p<0.01),除了蛋白质含量和碳水化合物含量,OTC与CK处理下其他生长指标在分化期差异均显著(p<0.05),并且OTC与CK处理下玉米叶片叶绿素a、b、碳水化合物、蛋白质含量在抽雄期差异均显著(p<0.05),也即增温处理促进了玉米的生长发育,并促成了地下生物量的累积,其中在成熟期的促进作用表现得最为明显。
2.1.3 叶片酶活性和丙二醛含量 由表3可知,模拟增温显著提高了玉米叶片超氧化物歧化酶、过氧化氢酶、过氧化物酶活性和丙二醛含量,在不同时期均表现为OTC>CK。随着生长期的进行,OTC和CK处理下玉米叶片超氧化物歧化酶、过氧化氢酶、过氧化物酶活性均呈逐渐增加的趋势,其中OTC与CK之间的差异随生长期的进行而逐渐拉大,在成熟期二者均达到极显著差异水平(p<0.01),OTC与CK处理在分化期差异均显著(p<0.05);而丙二醛含量随玉米生长期的进行呈逐渐降低趋势,二者在分化期差异极显著(p<0.01),在抽雄期差异显著(p<0.05),在成熟期差异不显著(P>0.05)。增温处理促进了玉米叶片超氧化物歧化酶、过氧化氢酶、过氧化物酶活性和丙二醛含量的提高,总体而言成熟期的促进作用表现得最为明显。

表2 模拟增温对玉米植株生理生化特性的影响

表3 模拟增温对玉米植株叶片酶活性丙二醛含量的影响
2.1.4 产量特性 由图1可知,模拟增温显著提高了玉米在成熟期的百粒质量、穗粒数和穗粒质量,与CK相比,分别增加5.3%、18.9%、8.2%,且穗粒数和穗粒质量均与 CK 之间差异极显著(p<0.01),百粒质量也表现为OTC显著高于CK(p<0.05)。由于百粒质量和穗粒数是玉米产量的构成因子,因此其增加将直接提高玉米产量,表明OTC在一定程度上促进了玉米穗粒数的增加,从而提高了玉米产量。

2.2 模拟增温对玉米根区土壤呼吸速率和酶活性的影响
2.2.1 土壤呼吸速率 由图2可知,模拟增温显著影响生长期玉米根区土壤呼吸速率。随着生育期的推进,OTC和CK玉米根区土壤呼吸速率呈先增加后降低趋势;同一时期玉米根区土壤呼吸速率均表现为OTC>CK,在分化期和抽雄期两者差异极显著(p<0.01),在成熟期两者差异不显著(P>0.05)。与CK相比,在分化期、抽雄期和成熟期,OTC处理玉米根区土壤呼吸速率分别提高15.6%、18.3%、3.5%。
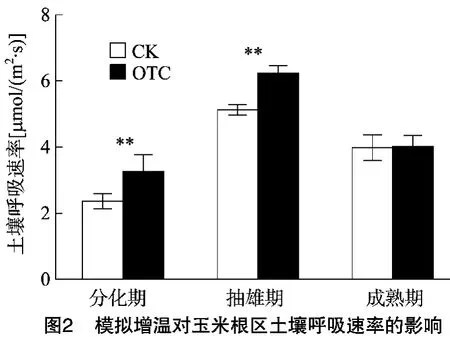
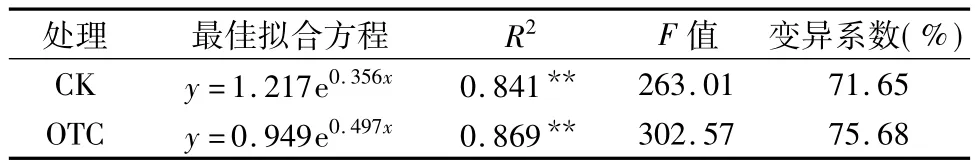
表4 玉米根区土壤呼吸速率(y)与土壤温度(x)的指数关系
由表4可知,在整个生长期,玉米根区土壤呼吸速率和土壤温度呈极显著的指数关系,CK玉米根区土壤呼吸速率(y)与土壤温度(x)的方程为:y=1.217e0.356x,OTC 玉米的方程为:y=0.949e0.497x,OTC和CK的指数回归方程可分别解释土壤呼吸速率71.65%和75.68%的变异。
2.2.2 土壤酶活性 由表5可知,模拟增温显著影响生长期玉米根区土壤酶活性。随生育时期的推进,OTC和CK玉米根区土壤酶活性呈先增加后降低的趋势,其中OTC与CK之间的差异随生长期的进行而逐渐拉大,同一生育时期,玉米根区土壤酶活性均表现为OTC>CK,在抽雄期差异极显著(p<0.01),在分化期差异显著(p<0.05),在成熟期差异相对较小。增温处理促进了玉米根区土壤酶活性,其中在抽雄期的促进作用表现得最为明显。

表5 模拟增温对玉米根区土壤酶活性的影响
由表6可知,在玉米的整个生长期,土壤酶活性与土壤温度呈极显著的指数关系,CK玉米根区土壤酶活性(y)与土壤温度(x)的方程为 y=1.012e0.269x,OTC 处理玉米根区土壤酶活性与土壤温度的方程为y=0.915e0.384x,CK和OTC的指数回归方程可分别解释土壤酶活性79.23%和83.24%的变异。
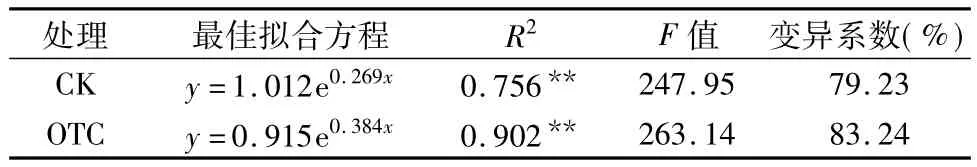
表6 玉米根区土壤酶活性(y)与土壤温度(x)的指数关系
3 讨论与结论
3.1 模拟增温对玉米生长特性的影响分析
开顶式生长室(OTC)是一种较为理想的试验升温设备,也是研究全球气候变暖的有效工具[1,14],在全球各种模拟增温的控制性试验已得到广泛的应用。本研究采用OTC进行小尺度的模拟控制试验研究玉米对增温的响应。受到OTC内温度的影响,OTC内土壤温度显著增加,土壤湿度显著降低,不仅玉米生长较快,叶面积大,分蘖数多,而且比叶重和叶面积指数均高于CK,该结果表明增温能够促进玉米植株的的生长和促进其产量的提高。在此过程中,主要是通过促进玉米的光合代谢能力、物质的合成与积累,从而提高产量,这与前人的研究结果[15-16]一致。玉米的产量主要是由光合作用产生的有机物质决定,而叶面积指数有利于捕获更多的太阳光能,本研究中,玉米对增温的响应较为敏感,同时,发达的根系和较小的R/S是实现其新陈代谢的稳定的基础,这也是玉米适应气候变化的重要生理机制[17-18]。而此过程中,增温也调控了土壤健康状况(提高了土壤酶活性和土壤呼吸速率),从而为玉米的生长提供了更好的条件。
3.2 增温对玉米生理指标的影响分析
能量和物质代谢是玉米生长发育的基础,温度调控各个生理、生化过程。由表2可知,随着生育期的推进,OTC和CK玉米叶片叶绿素含量、碳水化合物和蛋白质含量呈逐渐增加趋势,相同时期均表现为OTC>CK。增温主要通过影响玉米叶绿素含量而间接影响植物的光合生产能力,从而促进了叶绿素的合成。因此,增温对玉米叶片光合生理特性和叶绿素含量的变化保持一致。此外,增温处理能够增加玉米叶片碳水化合物、叶绿素和蛋白质含量,一方面说明玉米可能适应更高的增温环境,另一方面说明了碳水化合物和蛋白质作为植物体的基础性物质,参与了营养期干物质的合成与积累,为了适应环境改变而作出的积极的生理调节,对玉米的生长具有重要的生态学指示意义[18]。此外,在玉米生长的不同时期,增温对玉米均有一定的促进作用,尽管促进作用不尽一致,但总体表现为成熟期的促进作用最为明显,这与玉米自身调节营养物质的平衡有关,还有可能是土壤动物、微生物、植物根系或者非生物因子共同调控所产生的结果[19]。
3.3 增温对玉米土壤特性的影响分析
土壤酶活性能够催化土壤的生物化学循环,在一定程度上反映了生物化学过程的方向和强度。温度往往是影响土壤酶活性的重要因子,在全球气候变化背景下,土壤酶活性在生态系统的养分循环中起着重要的作用。本研究中,增温处理下玉米土壤呼吸和土壤酶活性均显著提高,与前人的研究结果[20-21]一致;本研究还发现,土壤呼吸和土壤酶活性均与土壤温度呈现出较好的一元指数关系,由此表明了增温对玉米土壤酶活性和土壤呼吸均有一定的促进作用。作为一种活性蛋白土壤酶和胞外酶,在一定的温度条件下,温度的升高有助于土壤酶活性和土壤呼吸的提高[22];此外,土壤温度的升高有助于提高养分的可吸收性、有效性,促进了土壤微生物群落的增殖,进而促进土壤生态系统的物质循环和能量流动[23-24]。
玉米的形态和生理特征对增温的响应机制至今仍存在很大的不确定性。本研究连续3年的控制性试验基于小尺度,未能完全反映出大尺度下的全球气候变暖对玉米的形态和生理特征的影响。值得注意的是,该试验背景下的光照、CO2通量、水分等微环境因子仍有待进一步观测和分析,未来应重点着手于长期观测和自然状态下的微环境因子的相结合。
