模拟干化土壤中的植被生长及土壤水分变化
2018-10-12白永红高志永汪有科董建国
白永红,高志永 ,陆 静,汪有科 ,董建国
(1.西北农林科技大学资源环境学院,陕西杨陵712100;2.西安理工大学西北旱区生态水科工程国家重点实验室培养基地,陕西西安710048;3.中国科学院教育部水土保持与生态环境研究中心,陕西杨陵712100;4.西北农林科技大学水利与建筑工程学院,陕西杨陵712100)
土壤干化是黄土高原地区存在的一种特殊水文现象,近50 a气候变暖和降雨减少是造成黄土高原地区土壤干化的主要原因,而植被类型选择不当,种植密度过大,生产力过高,大范围植被重建加速土壤水资源的消耗,导致土壤干化进一步加深[1],甚至形成永久性干层[2]。如 Y.Q.Wang[3]等发现黄土高原林地土壤干层厚度达最大,李军[4]等通过对23种林地土壤水分的分析发现土壤干层厚度达到或超过10 m。土壤干层的存在会影响土壤水分运移,阻碍降水入渗补给,弱化“土壤水库”功能,导致土壤质量和土壤生产能力降低,进而影响植被的水土保持、水文调节、水源涵养等生态系统服务功能[5-8]。
迄今,关于土壤干化研究主要集中在量化指标、影响因素、形成过程、模型预测和人为调控上[1-2,6,9-14]。土壤干化的形成,不仅制约已有植被生长,而且会对后续植被的选择和生存影响巨大[10,15]。目前关于该问题的研究较为薄弱,此外,尚未见对深层土壤干化后续栽植多年生植物进行报道。在黄土高原大规模退耕还林还草的背景下,研究干化土壤水分和再植植被生长的动态及其互馈关系对于维持该区域生态稳定至关重要。
本研究模拟生长28 a的山地苹果(Malus pumila)园伐后干化土壤,并在干旱土壤上栽植早熟禾(Poa pratensis)、柠条(Caragana korshinskii)、苜蓿(Medicago sativa)和刺槐(Robinia pseudoacacia)4种典型植物,利用CNC-503B型中子仪于2014-2016年监测4种典型植被0~10 m土壤水分,测定其植被株高、地径和生物量,以期对干化土壤后续植被建造及深入研究人工林耗水形成的干层治理提供参考。
1 研究区概况
研究区位于陕西省米脂县境内远志山红枣(Ziziphus jujuba)示范基地(37°12'N,109°28'E),为典型黄土高原丘陵沟壑区;属中温带半干旱性气候,年平均气温8.4℃,极端最高气温38.2℃,极端最低气温-25.5℃;2012-2015 年降雨量分别为 404.4、530.1、460.4 mm 和 334.8 mm,年均降水量 451.6 mm,主要集中在夏季,其中7、8月降雨量占全年降水量的49%;土壤以黄土母质发育的黄绵土为主,容重为1.24 g·cm-3,质地为粉质沙壤土。研究区1984-2007年间为旱作苹果(Malus pumila)园地,2007年苹果林伐后休闲至2011年栽植枣树,同年布设试验区。
2 研究方法
2.1 试验土柱设置
试验在一水平阶地上布设,试验土柱为12个直径0.8 m、深10 m的大型土柱。土柱采用开挖后回填,开挖时按照之前测定的土壤质地层分3层开挖,并将3层土壤分别堆放保存,以保证回填时按原来的土壤质地层次分层回填。土壤回填前用厚约1 mm的大棚塑料膜铺设在土柱井壁,使柱体土壤与外界土壤隔离,避免土柱内外水分交流扩散。回填时,一方面按照之前测定的土壤质地分层回填,逐层压实;另一方面重点控制回填土的土壤容重(1.29±0.05 g·cm-3)和含水率(7%左右),从而最大程度地模拟旱作枣林地干化土壤。回填土经过均匀掺混后分层回填踩实,土柱内垂向剖面土壤含水量较均一,含水率7.5%左右。12个土柱分别设置为早熟禾、柠条、苜蓿和刺槐的4个植被处理,每种处理3个重复。栽植时间是2011年4月。其中刺槐是栽植2年生苗木,早熟禾、苜蓿和柠条均为种子直播。
2.2 观测指标
2.2.1 土壤水分测定 在每个土柱的中间位置安置10 m长铝管,利用CNC-503B型中子土壤水分仪观测,试验于2014年5月至2016年10月每月观测3次,分别于每月3日、13日和23日测定,若遇雨雪天气则后延。
土层储水量(W)根据土壤体积含水率计算:

式中,ω为体积含水率(%);h为土层深度(cm)。
为了反映土壤水分的亏缺和补偿状况,采用土壤储水亏缺度和土壤储水亏缺补偿度[16]来描述4种典型植被干化再植土壤水分状况。
土壤储水亏缺度(%):

式中,Fw为凋萎含水量(mm);Wc为实际含水量(mm)。
土壤储水亏缺补偿度(%):

式中,We为年末土壤实际含水量(mm);Wb为年初土壤实际含水量(mm);Fc为田间持水量(mm)。
土壤水分在土层中运动状况,可以用方差来描述,方差>1的土层为活跃层,方差<1的土层为稳定层[17]。方差公式如下:
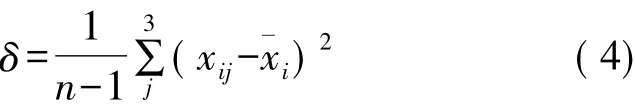
式中,xij为j年i层的土壤含水量(%);xi为土壤含水量在2014-2016年i土层均值(%)。
2.2.2 生物量测定 2015年开始测定全部试验土柱内的株高、密度(株·m-2)、地径(柠条),刺槐以土柱试验处理中的3棵林木为样本,每隔7 d测量其各个枝条的长度和基部直径。地上生物估算,每月一次采用在试验区附近收获与试验土柱内相同的植物、相同规格和数量的枝条和草叶,室内烘干,并用电子天平称重。
2.2.3 耗水量 试验区植被耗水量利用农田水量平衡法计算。试验区植被均为雨养,无灌水,不发生深层渗漏,无地下水补给,试验期间未发生地表径流。因此,植被耗水量(ET,m2·hm-2)公式可简化为:

式中,Pr为降雨量(mm);ΔW为计算时段初与计算时段末土壤储水量之差(mm)。
2.2.4 水分利用效率 水分利用效率(kg·m-3)指作物消耗单位水所制造的干物质量,公式为:

式中:Y为干物质量(kg·hm-2)。
2.3 数据处理
用SPSS18.0软件进行数据统计分析,利用origin2016绘图软件作图。
3 结果与分析
3.1 不同植被下土壤水分变化
图1为早熟禾、苜蓿、柠条和刺槐在月和季尺度上土壤含水率的变化,4种植被土壤含水率在1-4月及11-12月维持相对稳定,在5-10月受降雨及植被耗水等影响,波动较大;在季尺度上,土壤含水率变化在冬季较低,其他季节相对稳定。4种植被种植初期及2014-2016年不同土壤深度处水分状况表明(图2),早熟禾、柠条、苜蓿和刺槐在种植初期0~10 m土层土壤含水率相近,分别为7.44%、7.47%、7.31%和 7.50%。除早熟禾外,柠条、苜蓿和刺槐种植地0~10 m土层土壤含水量于2014年后低于初始含水量;与初始含水量相比,早熟禾种植地土壤含水量变化率为 0.46%、0.61%和 1.14%;柠条种植地土壤含水率变化量为 0.62%、-0.40%和-0.55%;苜蓿种植地土壤含水率变化量为0.65%、-0.29%和-0.62%;刺槐种植地土壤含水率变化量为0.28%、-2.29%和-2.30%。从作物耗水量来看(表1),早熟禾耗水量低于同期降水量,其耗水量在2015年和2016年均显著低于同期其他植物的耗水量(P<0.05),是柠条、苜蓿和刺槐的0.78~0.89、0.78~0.86 倍和 0.53~0.91 倍;柠条、苜蓿和刺槐耗水量在2015年和2016年高于同期降水量;刺槐耗水量在2014年和2015年显著高于其他(P<0.05),其耗水量是同期(2014年、2015年)降水量的0.94倍和1.79倍;苜蓿耗水量在2016年显著高于其他(P<0.05),是同期(2016年)降水量的1.06倍。

图1 在月和季尺度上早熟禾、苜蓿、柠条和刺槐地土壤含水率Fig.1 Soil water content in pratensis,caragana,alfalfa and robinia on the monthly and seasonal scales

图2 早熟禾、苜蓿、柠条和刺槐土壤含水率Fig.2 Soil water content in pratensis,caragana,alfalfa and robinia

表1 不同植被耗水量Table 1 Water consumption of Pratensis,caragana,alfalfa and robinia
图3为4种植被种植地土壤含水率的方差分析。就活跃层而言(表2),早熟禾、柠条、苜蓿和刺槐地土壤水分活跃层深度分别为 2.6、3.2、3.0 m 和10 m。早熟禾土壤水分不存在亏缺现象(DSW<0),得到降雨的补充(CSW>0);早熟禾土壤水分均值显著高于其他3种植被土壤水分(P<0.05),其土壤水分在2015年(枯水年)达9.53%,是其他3种植被的1.57、1.51 倍和 1.83 倍,其土壤水分在 2016 年(丰水年)达 11.54%,是其他 3 种植被的1.96、1.90 倍和2.23倍。柠条、苜蓿和刺槐土壤水分在2014年后出现亏缺状态(DSW>0),降雨无法补偿亏缺的土壤水分(CSW<0),使土壤水分亏缺呈现逐年增加趋势,其中刺槐土壤水分亏缺显著高于苜蓿和柠条(P<0.05),DSW值>17%;刺槐土壤水分均值在2015年(枯水年)和2016 年(丰水年)分别为5.2%和5.18%,这表明刺槐在枯水年(2015)已耗尽土壤中可用的土壤水分,即使在丰水年(2016),亏缺的土壤水分也得不到补偿,只能依靠当年降雨维持生长;此外,柠条和苜蓿土壤水分亏缺程度无显著差异(P>0.05)。就稳定层而言(表2),早熟禾、苜蓿和柠条土壤水分接近初始含水量,土壤水分均值无显著差异(P>0.05),其范围为 7.04%~7.58%;早熟禾土壤水分无论在枯水年(2015)还是丰水年(2016)向下移动,出现显著补偿稳定层土壤现象(CSW>0,P<0.05),而柠条和苜蓿稳定层土壤水分得不到补偿(CSW<0)。
3.2 模拟干化土壤中的植被生长
模拟干化土壤中早熟禾、柠条、苜蓿、刺槐各处理生育期内株高、地径、单株生物量见图4和表2。图4表明,植被株高、地径和单株生物量均随着时间增长,达到一定值后趋于稳定。由表3可以看出,4种植被的株高生长量存在显著性差异(P<0.05),其中早熟禾株高生长量最小,为17.6 cm和22.7 cm,刺槐株高生长量最大,为53 cm和55.5 cm;地径的生长量也存在显著性差异(P<0.05),其中苜蓿地径变化量最小,为1 mm和1.2 mm,刺槐地径变化量最大,为 3.6 mm 和 3.9 mm;早熟禾、柠条、苜蓿单株生物量无显著差异(P>0.05),刺槐单株生物量显著高于其他三者(P<0.05),是上述3种植被生物量的 22.7~33.5倍。此外,同种植被株高生长量和单株生物量在丰水年(2016)显著高于枯水年(2015,P<0.05),地径生长量在丰水年和枯水年无显著变化(P<0.05)。

图3 不同植物土壤水分方差分析Fig.3 Variance of soil moisture of different vegetation types
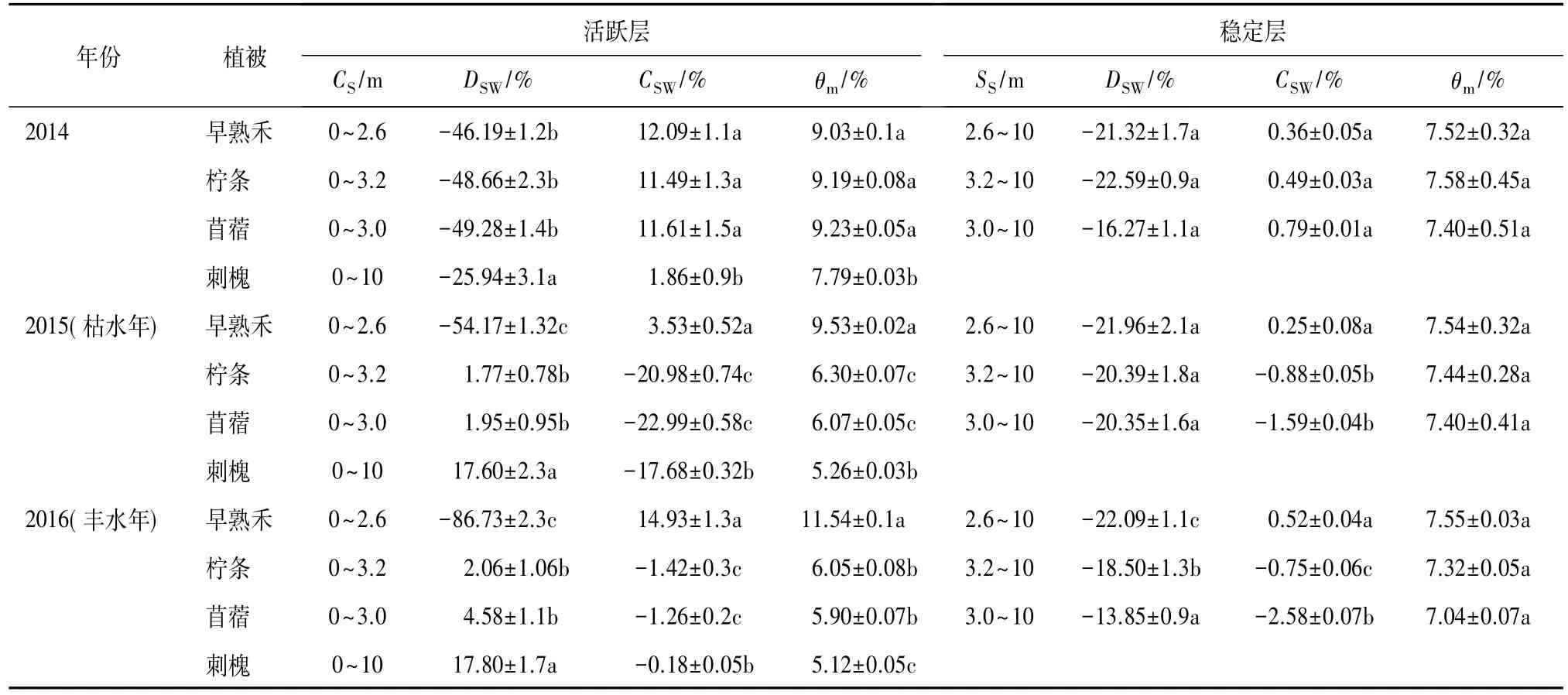
表2 不同植物土壤水分在活跃和稳定层亏缺和补偿状况Table 2 Soil water deficit and compensation in changeable and stable layers for different vegetation
3.3 不同植被水分利用效率
表4为2015-2016年干化土壤中不同植被生物量及水分利用效率。可以看出,早熟禾的生物量显著低于其他植被(P<0.05),其生物量分别是柠条、苜蓿和刺槐的 0.6、0.8、0.4 倍(2015 年)和 0.6、0.9、0.6倍(2016年)倍;刺槐生物量显著高于早熟禾,分别达 21 100 kg·hm-2和 24 100 kg·hm-2;早熟禾和苜蓿的水分利用效率无显著差异(P>0.05),该值的范围为 2.6~2.9 kg·m-3;柠条的水分利用效率显著高于早熟禾和苜蓿(P<0.05),是早熟禾和苜蓿的1.2倍和1.5倍。此外,模拟干化土壤中4种植被生物量受当年降水量影响较大,丰水年(2016)生物量高于枯水年(2015),其中柠条生物量变化最大,丰水年生物量是枯水年的1.6倍;早熟禾和苜蓿的水分利用效率在枯水年高于丰水年0.2 kg·m-3和0.3 kg·m-3,而柠条和刺槐的水分利用效率丰水年高于枯水年,刺槐在丰水年的水分利用效率是枯水年的1.7倍。
4 结论与讨论
4.1 结论
通过在模拟干化土壤中种植植被并监测土壤水分及各生长指标,分析各植被土壤水分动态变化、植物生长状况及水分利用效率,得到以下结论:

图4 2015年和2016年不同植被株高(a、b)、地径(c、d)及生物量(e、f)Fig.4 Plant hight(a,b),ground diameter(c,d)and individual biomass(e,f)for different vegetation in 2015 and 2016
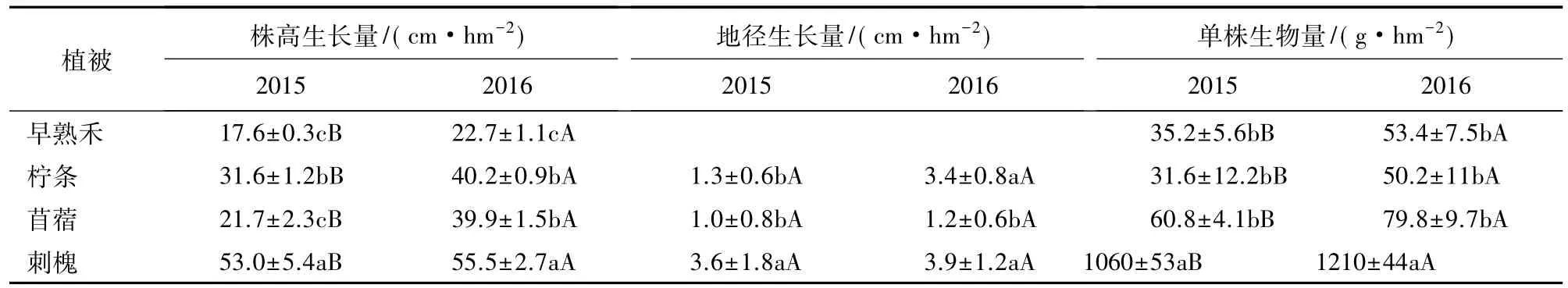
表3 不同植被生长情况Table 3 Growth of different vegetation
注:不同大写字母表示同一植被在不同年份同一处理存在显著性差异(P<0.05)。

表4 不同植被2015-2016年生物量、耗水量及水分利用效率Table 4 Biomass,ET,WUE of different vegetation in 2015 and 2016
1)干化再植早熟禾在监测期(2014-2016年)年耗水量均<当年降水量,土壤储水量及水分变化深度逐年增加,使得土壤水分活跃层(0~260 cm)的水分得到恢复,该层次土壤的储水量较初期增加115.07 mm。
2)干化再植柠条和苜蓿在2014年耗水量<当年降水量,其 0~320 cm(柠条地)和0~300 cm(苜蓿地)土层土壤水分(活跃层)得到临时补充,该土层储水量较初期分别增加57.42 mm和54.28 mm,逢枯水年(2015)该土层土壤水分消耗殆尽,再次出现干化现象,遇丰水年(2016)此层土壤水分得不到补偿。
3)干化再植刺槐在2014年耗水量<当年降水量,活跃层(0~1 000 cm)土壤水分变化剧烈,2014年后干化加剧,至2016年0~1 000 cm范围内土壤储水量较初期减少241.57 mm。
4)模拟干化土壤中种植早熟禾、苜蓿、柠条和刺槐,其生长受当年降水量影响较大,株高生长量和单株生物量在丰水年(2016)显著>枯水年(2015)。
4.2 讨论
土壤水分是限制黄土高原植被生长的关键性因子[18],黄土高原土壤干化日趋严重,干层空间分布增大[8]。植被选择不当、种植密度过高等因素促进干层进一步恶化[19]。为分析干化条件下土壤水分及植被生长状况,于2011年在土壤干化的基础上栽植早熟禾、柠条、苜蓿和刺槐4种典型植被,研究表明,干旱土壤再植早熟禾能够恢复0~10 m土层土壤水分;干化再植柠条和苜蓿初期在降雨量充沛条件下(2011-2014年年均降雨量为495 mm)能够使土壤活跃层(0~3 m)土壤水分得到临时恢复,后期遇枯水年(年降雨量342 mm)土壤活跃层再次出现干化。已有研究表明早熟禾水分利用层和87%的主根系分布在0~30 cm土层内[20],柠条和苜蓿在干旱胁迫下主根生长受限,生物量下降,减弱“大根系”冗余消耗过多的水分和养分,使其趋向适应干化环境,显著降低蒸腾耗水[21-22],监测期降雨量(2011-2014年)不仅能满足植被生长而且能够入渗至2~3 m补充土壤水分[23],至2014年活跃层土壤水分在降雨的补给下达到9.03%~9.23%(表2),较初期干化土壤水分增加1.8%~1.92%。在枯水年(2015年),早熟禾仅消耗0~30 cm表层土壤水分,其耗水量低于同期降雨量,部分降雨入渗至较深土层,而柠条和苜蓿的耗水量是降雨量的1.3倍和1.2倍,降雨无法满足柠条和苜蓿的需求,它们只能过度消耗活跃层土壤水分来满足生长,最终造成活跃层土壤再次干化,致使活跃层土壤水分由原来的9.13%,9.23%降至 6.3%,6.07%,稳定层(3~10 m)干化难以得到降水补给,土壤水分长期维持在初期干化状态(7.4%),即使遇丰水年(2016年),活跃层和稳定层的土壤水分并不能得到恢复。柠条林地11年后2~9 m土层全部干化[24],苜蓿地7年后土壤干层达到10 m[25]。预计后期,柠条和苜蓿耗水深度还会增加,稳定层土壤水分会进一步被消耗,再次出现通体干化现象。如果要实现干层土壤水分恢复,只能通过撂荒或种植如早熟禾等浅根系1年生植被来降低耗水强度,经多年降雨入渗,逐步恢复活跃层土壤水分,通过较长时间恢复稳定层土壤水分。
干化再植刺槐土壤水分活跃层0~10 m,即0~10 m土层土壤水分变化剧烈,这与已有的研究结果一致[26],随着林龄增加,土壤干旱化加重,2011年初始干化土壤含水率为7.50%,2016年降至5.18%,也就是说干化再植刺槐不仅达不到恢复土壤水分的目的,而且使得土壤干化进一步加重。刺槐是干旱敏感型树种,对水分的需求量较高,林地土壤水分多寡影响蒸腾耗水大小[27],刺槐生长前期,在降水和土壤水分供给条件下,生长旺盛,蒸腾耗水大于降雨量,使得深层土壤水分降低,在2015年(枯水年)将前期储存在土壤中水量消耗殆尽,在2016年(丰水年)只能依靠当年降雨维持生长,其耗水量与降雨量接近。随着林龄的增长,其生产力将会明显降低且年际波动剧烈,最终出现生长衰败和死亡现象。此外,刺槐林地干化程度和干化发展速度与造林密度有关,密度过大,出现干化程度越严重,干化发展速度越快,生长衰败越早[26]。
植物的水分利用效率反映了植物在消耗水分后所固定CO2(干物质积累)的量,可反映植物生长与水分利用的关系,高的水分利用效率被认为植物在干旱半干旱地区成功生长的一个特征[28-30]。早熟禾对土壤水分消耗方式属于迅速消耗型,土壤含水量充足时蒸腾剧烈[31],其在2016年活跃层土壤含水率达11.54%,较 2015年高 2.01%,造成 2016年耗水是2015年的1.62倍,尽管生物量2016年高于2015年,但高耗水导致其水分利用效率在丰水年降低。苜蓿、柠条和刺槐的水分利用效率随着干旱胁迫的加剧而升高,土壤水分的亏缺降低植物的气孔导度,为了维持光合速率,叶片胞间CO2浓度和大气CO2浓度比值降低,提高了水分利用效率[32]。然而植物的水分利用效率对干旱胁迫相应的存在阈值,如果干旱胁迫进一步加剧,气孔表皮细胞失水过快,光合系统受损,气孔被动开大,降低了气孔限制,导致光合速率降低,蒸腾速率升高,促使水分利用效率降低[33]。如苜蓿活跃层土壤水分降低0.17%,水分利用效率降低了0.3 kg·m-3,刺槐活跃层土壤水分降低 0.14%,水分利用效率增加了 1.7 kg·m-3,即不同植被对干旱胁迫阈值不同,苜蓿干旱胁迫超出阈值,水分利用效率降低,而刺槐还在阈值范围之内。此外,干化再植植被受制于干旱胁迫,其生长量和生物量随着降水量的年际变化发生波动性变化,丰水年份生长量>枯水年份。
