修剪高度对季节性温度变化下早熟禾生理生态特性的影响
2018-09-19宋娅丽王克勤
宋娅丽, 王克勤
(西南林业大学生态与水土保持学院, 云南 昆明 650224)
由于城市扩张和人们对草坪生态系统的广泛认可,加之种植草坪会对环境、大众审美以及娱乐等方面带来诸多益处,全世界范围内总的草坪覆盖面积仍会不断增加[1]。修剪是草坪主要的管理措施之一,合理的修剪方式是维持草坪草再生能力的重要保障。修剪可能会导致植物损失大部分生长组织和光合器官,限制植物的生长;但同时又会刺激侧枝生长和新生组织再生,改变植株生物量的空间分配或净光合效率[2]。冷季型草坪草地上部分生长的最适温度介于16~24℃之间[3],修剪高度的差异会直接影响植物的光合作用[4],从而通过改变草坪地上部分生物量和颜色来影响草坪质量。同时,修剪高度还会通过改变草坪收获太阳能的能力来调节供植物生长的碳水化合物的量,但因植被类型不同而结果各异,主要表现为修剪高度改变会导致碳水化合物增加、减小或不变[5-7]。
草坪草的生长取决于光合作用吸收的CO2量,以及地上部分、根系和土壤通过呼吸作用释放的CO2量,这个平衡会随着季节性温度变化和植被或土壤条件变化而发生改变[8]。除自然条件外,适宜的修剪高度既与种植地区的气候、土壤等生态条件有关,也受修剪强度、栽培管理水平和植被类型的影响[4,9]。目前对于人为管理的草坪系统,研究集中于草坪生物量、土壤有机碳固定、碳吸收容量等方面[10-12]。但对季节性温度变化下修剪高度是否会影响成熟草坪草的生长,以及是否会补偿高温对冷季型草坪草生长带来的不利影响尚不清楚。因此,本研究通过连续两年(2012—2013年)田间试验,测定早熟禾(Poapratensis‘Baron’)生长季温度变化下修剪高度对成熟早熟禾的草坪外观质量、地上部分生物量、光合作用、呼吸作用和碳水化合物等指标,分析修剪高度对季节性温度变化下早熟禾生理生态特性的影响,旨在为冷季型草坪草可持续利用、生态保护与固碳增汇等多元化经营提供必要理论依据和实践基础。
1 材料与方法
1.1 植物材料和生长条件
试验地设置在美国新泽西州立大学(Rutgers)草坪研究基地(40.470oN~74.426 °W),试验时间为2012年和2013年的5—10月。该地区为大陆性气候,1月平均气温为-2~3℃,7月平均气温为21~24℃,年降水量为1 000~1 200 mm。植物材料为成熟(8年生)的草地早熟禾(Poapratensis‘Baron’),土壤类型为尼克松沙壤土(含41%沙土、39%泥土和20%粘土)。
1.2 试验设计
在成熟草地早熟禾草坪中,设置两块面积分别为2.5 m×3.0 m的样地,在两个样地内分别设置4个重复样地。试验以3.8 cm的修剪高度为对照(家庭、公园的修剪保留高度为3.8 cm),在两块样地修剪高度分别为7.6 cm(高修剪高度,HM)或3.8 cm(低修剪高度,LM)。每两周修剪一次;每周灌溉两次,通过灌溉的方式调节土壤含水量,使其在测定以下各指标前保持在20%的野外田间土壤含水量。为了使草坪正常生长,分别在2012年和2013年春季4月,施复合肥料(N、P、K的比例为16:4:8)一次,在6月中旬再施一次浓度为3.2 g N m-2的肥料。为了避免交叉影响,在整个培养过程中不使用杀虫剂。2012年和2013年的5—10月的日温度和光照辐射水平由坎贝尔科学气象站在Mesonet网络(40.470° N,-74.426° W)上获得。
1.3 气体交换参数测定
在2012年和2013年5月—10月每月的第一周,选择晴天,使用便携式红外线分析仪(LI6400,LI-COR Inc.,Lincoln,NE,USA)测定日照下的CO2气体交换量,包括草坪系统光合作用(Pn)和呼吸作用(Rtotal,包括地上部分、根系和土壤的总呼吸作用)。系统Pn和Rtotal根据Lohila等[6]描述的方法来测定。每年生长季开始(4月),每月将直径为10 cm的钢圈(高度为10 cm)放置在4个重复样地的土壤中,每个重复样地各5个,土壤高度为5 cm。在两个不同修剪高度下,气体交换量分别在20个重复土壤环(直径为10 cm)下测定。在测定Pn和Rtotal的同时,测定光照条件下的日平均温度,用于以下各结果图中Pn和Rtotal测定时的温度。
1.4 生理指标测定
在每月测定气体交换量的同时,测定草坪外观质量(TQ)。TQ是总的草坪性能的指示器,基于2012年和2013年5月—10月早熟禾地上部分密度、均一性和颜色按照1到9的数值来评价,1是最低、完全枯萎和棕色的草坪,而9是长势好、水分充足和颜色鲜绿的草坪[3,13]。每月在测定完植被光合作用和系统呼吸作用后,将气室面积内的地上部分从土壤中切断。4个重复样地中,5个重复土壤环中的早熟禾地上部分生物量(AB)带回实验室,放置在-80℃液氮中,冻干后称地上部分干重。非结构性碳水化合物(TNC)含量根据Ting[14]描述的修订方法来测定。
1.5 数据分析
不同修剪高度下的每个重复样品,根据国际通用统计软件SAS(SAS 9.1;SAS Institute Inc.,Cary,NC)中线性回归模型分析修剪高度、温度对植物生长的影响(ANOVA)。采用方差分析确定不同修剪高度对温度变化下TQ、SB、Pn、Rtotal和TNC上的差异显著性。计算各个指标的平均值和标准差,进行方差齐性检验和F检验,当F检验显著时,在95%的置信区间下进行LSD检验,同时计算LSD值。
2 结果与分析
2.1 生长季大气温度和光照辐射变化特征
图1为2012年和2013年生长季中早熟禾日最大温度和日照辐射月平均值。实验中每块样地有4个小样地,每个小样地有5个土壤环,每月在两个高度的样地中各测定20个重复,共40个。结果图中横坐标轴温度为日均温,测定中日均温共计42个(每个月测定7天,共6个月),剔除相同温度值后,每年得到18个不同温度。下文结果中草坪质量、地上部分生物量、光合作用、呼吸作用均是在日均温下的变化值,各个值均是每天20个重复值的加权平均值。每月测定光合作用和地上部分生物量前,均在不同样地中各设置20个土壤环。
由图1可知,两年中随着季节性温度变化,5月、6月、9月和10月的平均大气温度在冷季草生长的最适温度范围(10~24℃)。但在2012年夏季7月和8月温度平均值分别为32.6℃和30.7℃,2013年分别为29.4℃和26.0℃,均高于最适温度的范围。平均日照辐射水平与日最大温度变化一致,从5月到6月逐步增加,并在夏季7月和8月达到最高值,秋季9月和10月又随着时间的持续而有所下降。2013年最大温度和日照辐射略低于2012年。本试验是在两年的整个生长季中(5月到10月)测定随着温度的变化早熟禾生长和碳交换量的变化。
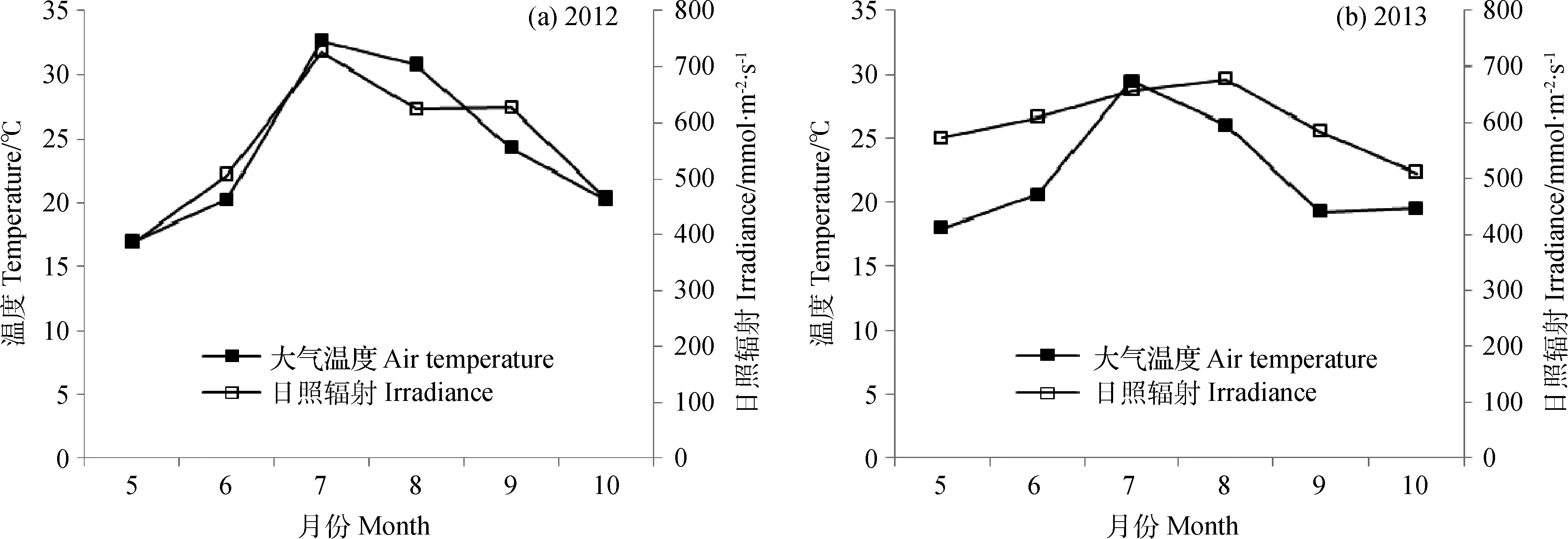
图1 2012年(a)和2013年(b)5—10月生长季中日最大温度、日照辐射特征Fig. 1 Daily maximum temperature and solar irradiance during the periods of growth from May to Octoberduring 2012 (a) and 2013 (b)
2.2 修剪高度对温度变化下早熟禾生长的影响
从图2可以看出,两年中在高修剪高度(HM)和低修剪高度(LM)下,早熟禾草坪外观质量(TQ)从5月到10月随着温度的上升直到21℃(2012年)和22℃(2013年)而逐渐平稳的增加;随后随着温度的上升而逐渐减小,当温度最高时,TQ达到最低值。2012年生长季,LM下TQ在26℃降低到可接受的最小值(6.0),而HM下温度直到上升到28℃时TQ才降低到6.0(图2a)。在两种修剪高度下温度均在26 ℃时,TQ降低到可接受的最小值,2012年26℃下的HM和LM分别比21℃下降26%和63%,2013年26℃下的HM和LM分别比22℃下下降78%和110%,在LM下的下降幅度更显著(图2b)。两年生长季中,温度分别在12~35℃和13~33℃时,HM下的TQ显著高于LM;在两年中当温度高于24℃时,不同修剪高度下TQ的差异更显著。

图2 2012年(a)和2013年(b)在两种修剪高度下温度变化对草坪质量(TQ)的影响。Fig.2 Effects of mowing height on turf quality (TQ) under different temperatures in 2012 (a) and 2013 (b)注:图中不同小写字母表示不同修剪高度间差异显著(P<0.05),下同Note:Different lower case letters indicate significant difference among different mowing height at the 0.05 level. The same as below
2012年生长季大气温度变化范围较大,最低温度为12℃,最高温度上升至35℃(图3a)。当温度从12oC增加到23oC时,早熟禾地上部分生物量(AB)在HM和LM下分别提高了19%和10%。而当温度从23℃升高至35℃,HM和LM下的AB分别降低了30%和43%。在不同温度下HM的AB均高于LM。2013年温度变化范围较2012年较小,范围在13~33℃(图3b)。当温度从13℃增加到22℃,早熟禾AB在HM下表现出极小的增加,而在LM下的AB增加了45.3%。当温度从23℃增加到33oC,HM和LM下的AB分别降低了26.7%和39.1%。两年中HM下的早熟禾AB比LM下的高出40%~45%。总的来说,温度在22~23℃范围内时,AB无论在HM或在LM下均为最高,随后随着温度的增加而降低,直到温度上升至33℃或35℃,AB也降到最低。

图3 2012年(a)和2013年(b)在两种修剪高度下温度变化对早熟禾地上部分生物量的影响Fig.3 Effects of mowing height on shoot biomass under different temperatures in 2012 (a) and 2013 (b)
2.3 修剪高度对温度变化下早熟禾植被光合作用(Pn)和总呼吸作用(Rtotal)的影响
在2012年,当温度从12℃上升到24℃,HM和LM下的早熟禾Pn增加到最高值,随后当温度从25℃增加到35℃,Pn又逐渐降低(图4a)。温度从12℃增加到24℃,Pn在HM和LM下分别增加了109%和203%;当温度从24℃上升到35℃,早熟禾Pn在HM和LM下分别降低了55%和49%。HM下的Pn在不同温度条件下均高于LM下,高出12%~128%。在2013年,当温度从13℃上升到24℃,早熟禾Pn在HM下增加了28%;当温度从13℃上升到22℃,早熟禾Pn在LM下增加了98%(图4b)。当温度从24℃增加到33℃,Pn在HM下降低了30%;而当温度从22℃增加到33℃时,Pn在LM下降低了48%。在生长季中的所有温度梯度下,Pn在HM下均高于LM下,高出18%~121%。
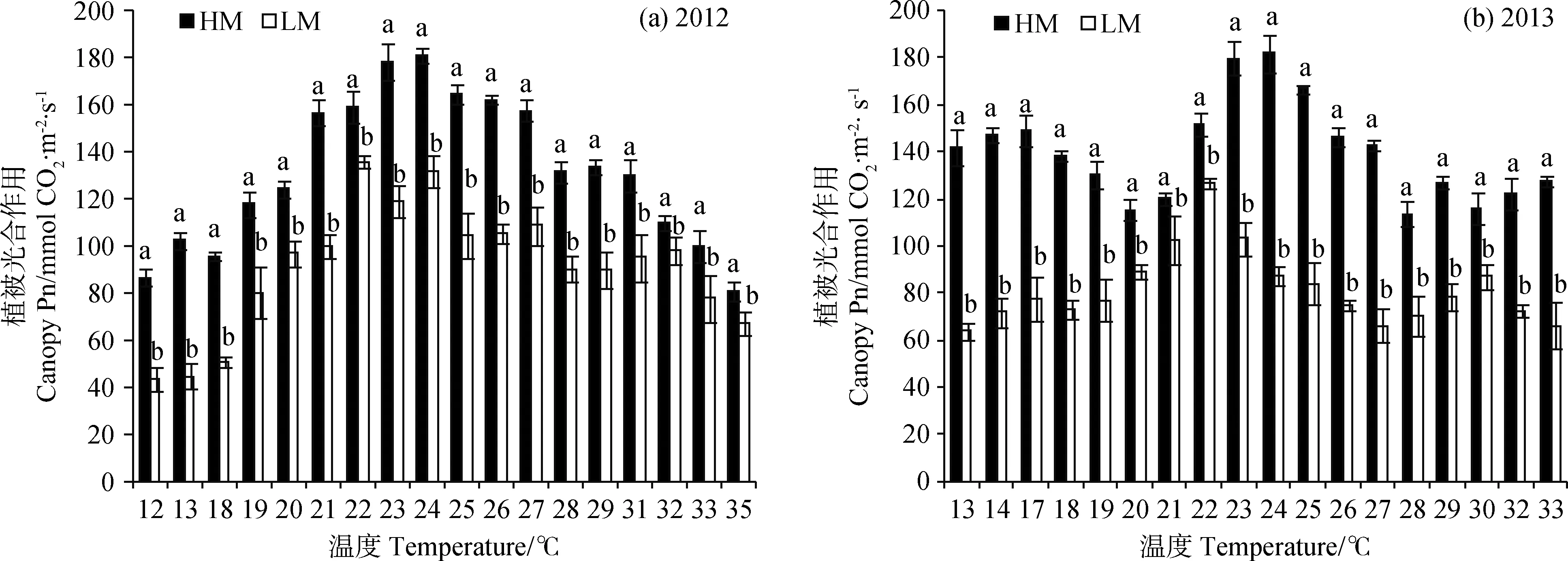
图4 2012年(a)和2013年(b)在两种修剪高度下温度变化对早熟禾植被光合作用(Pn)的影响Fig.4 Effects of mowing height on canopy photosynthesis (Pn) under different temperatures in 2012 (a) and 2013 (b)
图5为2012年和2013年在两种修剪高度下温度变化对早熟禾总呼吸作用(Rtotal)(包括地上部分、根系和土壤呼吸)的影响。从图中可以看出,两年中随着温度的增加,Rtotal在HM和LM下均表现出增加的趋势,但在不同的温度下植被总呼吸作用出现波动情况。2012年和2013年生长季中,当温度分别增加至或高于29℃和28℃,Rtotal值在HM下超过了Pn值。而在两年中的LM处理下,当温度分别增加至或高于29℃和25℃,Rtotal值超过了Pn值。
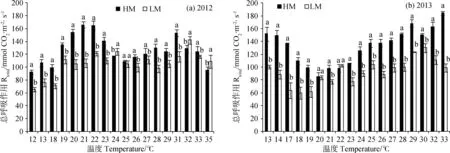
图5 2012年(a)和2013年(b)在两种修剪高度下温度变化对早熟禾总呼吸作用(Rtotal)(包括地上部分、根系和土壤)的影响Fig.5 Effects of mowing height on respiration rate of shoots,roots,and soil (Rtotal) under different temperatures in 2012 (a) and 2013 (b)
2.4 修剪高度对温度变化下早熟禾碳水化合物的影响
两年中随着温度的增加,早熟禾TNC分别在23~24℃和22~23℃下最高(图6),当温度高于或低于这个范围,TNC都有不同程度的减小。这与碳平衡最高值的温度范围是一致的。2012年和2013年温度分别从12℃增加到24℃以及从13℃增加到23℃,TNC在HM或LM下均表现出增加的趋势。随后随着温度的增加,TNC在两年中分别逐渐降低。当温度增加至32℃时,TNC在两年中的HM或LM下无显著差异。
3 讨论
管理措施和环境因素对草坪草形态学上的影响主要包括草坪生物量的改变、分蘖数和叶片高度的变化,以及颜色和叶片长度的改变等[15]。修剪高度是田间管理草坪草的典型措施,对草坪质量影响较大。本研究中早熟禾草坪外观质量随着修剪高度的降低而降低,与Salaiz等[15]在匍匐翦股颖(AgrostisstoloniferaL.)中的研究结果相似,修剪高度高的草坪质量较高,且颜色鲜绿。而相反地,Richie等[17]在高羊茅(FestucaarumdinaceaSchreb.)中的研究表明,修剪高度较低的草坪质量较高。Su等[18]在同一块样地对不同草坪做了修剪高度对草坪质量的影响研究,结果表明:在混播草坪(早熟禾PoapratensisL.×德州蓝草PoaarachniferaTorr.)中草坪质量随着修剪高度的降低而降低,而早熟禾(Apollo)的草坪质量随着修剪高度的降低而增加。另外,修剪高度和刀片锋利程度也会影响草坪整齐度、颜色以及草坪质量[19]。

图6 2012年(a)和2013年(b)在两种修剪高度下温度变化对早熟禾植被总碳水化合物(TNC)的影响Fig.6 Effects of mowing height on total non-structure carbon under different temperatures in 2012 (a) and 2013 (b)
生长季中温度的改变显著影响着草坪草生理生态指标,这表现在多种草坪草中[3,13]。例如控制环境实验中,匍匐翦股颖的光合作用最高值出现在20~24℃的范围内[20]。但是,冷季草植被Pn的最适温度是有变化的,而自然条件下早熟禾的最适温度并没有报道过。本研究中,两年生长季的HM和LM下,温度在22~24℃时植被的光合作用最高,说明这是野外管理条件下早熟禾的最适温度范围。无论在HM还是LM下,最高TQ和AB的最适温度在20℃和23oC之间。两年中,当温度超过最适温度时,TQ、AB和Pn随着温度的增加而降低;当温度分别升高到33oC或35oC的最高值时,以上各指标均降低到最低值。较低的修剪高度LM加剧了早熟禾TQ、AB和Pn在高温下的降低程度;但是LM并没有改变这些指标下的最适温度。
植物Pn和TNC是能量生产和可利用性的有效指标[21]。当植物碳水化合物产量高于消耗量,会形成正的碳平衡,正的碳平衡可以帮助植物最大限度的应对高温带来的压力。较低的修剪高度下,光的可利用性降低,绿叶数降低,降低地上部分的光合作用;同时,用来维持呼吸的非光合物质提高了呼吸作用,导致植物中能量的储存和可利用性降低。研究期间,在不同修剪高度下,早熟禾总碳水化合物随着季节性温度变化表现出很大的差异。在两年的试验中,2012年当温度<21℃和>33℃以及2013年当温度<22℃和>33℃时,TNC含量在LM下高于HM下的含量。TNC含量在低修剪高度下较高,可能是由于在这些温度范围内植被生长所需的碳水化合物有所下降。
在最适温度范围内,高修剪高度下的TNC含量高于低修剪高度。同样,Howieson和Christians[22]以及Peterson等[23]分别在温室中测定出匍匐翦股颖(AgrostisstoloniferaL.)和野牛草(Buchlo?dactyloides(Nutt.)Engelm.)碳水化合物积累量在较低的修剪高度下显著降低,可能由于植被种类和环境条件影响TNC含量[24-25]。在2年中,2种修剪方式下,TNC含量随着温度的增加而迅速增加,在24℃达到最高值,随后随着温度的增加而减小。说明TNC含量最高时的最适温度与地上部分生长的最适温度是一致的。Qian和Fu[26]也指出,低修剪高度(6.4 mm)下的匍匐翦股颖(L-93)与高修剪高度(25.4 mm)相比,TNC降低了32%~39%。植被中TNC在高温下的降低可能是碳重新分配或重新储存的结果。
系统碳收支取决于植物光合碳吸收和呼吸碳损失引起的碳平衡[27-28]。系统的碳汇能力主要取决于植物叶、根系的转化,这些转化产物会成为永久性的土壤碳库。本研究中早熟禾是否吸收和损失碳取决于大气温度,同时也会被割草高度所影响。2012年和2013年,当温度分别在20~23℃和17~27℃的范围内,早熟禾光合作用高于呼吸作用,说明在整个系统碳交换中,固定的碳高于损失的碳,在生态系统碳平衡上表现出碳增益。当温度高于植物生长的最适温度,植被在低割草高度下光合作用小于呼吸作用,光合作用与呼吸作用的差值变为负值,而这个变为负值时的温度低于高割草高度下的温度。另外,在LM下的大多温度中,光合作用小于呼吸作用;当温度低于或高于最适温度范围时,光合作用同样小于呼吸作用。而在HM的多数温度下,光合作用高于呼吸作用。这个结果说明草坪草在修剪高度低的情况下削弱了草坪的固碳能力,但加剧了碳损失潜力,形成负的碳平衡。同时,草坪草系统碳平衡对土壤和植物水分情况以及对修剪高度、季节性温度变化交互作用的响应,还有待进一步研究。
4 结论
在田间试验条件下,高修剪高度(HM)和低修剪高度(LM)下的早熟禾Pn最高值出现的温度范围在22~24℃,TQ和AB最高值的温度在20oC和23oC之间。当温度超过最适温度或在较低的修剪高度下,植物TQ、AB和Pn均有所降低,在33~35℃降低到最低值。整个草坪系统Rtotal随着温度的增加而增加,而当温度高于24℃时植被Pn降低,Pn小于Rtotal。在较冷的季节,早熟禾Pn高于Rtotal保持正的碳汇,而在夏季高温下随着温度的增加形成碳损失,导致使植物TQ和AB下降。修剪高度的增加可能会缓解高温对植物造成的不利影响,并在草坪系统内保持正的碳平衡。
