豆粕替代鱼粉对点带石斑鱼生长、消化和抗氧化能力的影响
2018-09-07安贸麟王庆奎孙金辉徐大为郭永军程镇燕
安贸麟, 范 泽, 王庆奎, 孙金辉, 徐大为, 郭永军, 程镇燕
(天津农学院水产学院/天津市水产生态及养殖重点试验室,天津西青 300384)
点带石斑鱼(Epinepheluscoioiaes)蛋白质含量高、脂肪含量低,不仅包含人体多种代谢所必需的氨基酸,而且富含多种无机盐和铁、钙、磷这些微量元素以及各种维生素。近年来,石斑鱼的养殖已经成为我国东南沿海,以及台湾东南部重要的海水养殖的水产品之一。但随养殖规模的日益扩大,对鱼粉的需求量也不断增大,而全球鱼粉的年产量却日趋下降,其价格一直处在高位,使饲料成本居高不下[1]。因此,寻找到节约饲料鱼粉用量的适宜途径成为点带石斑鱼配合饲料开发的研究热点及难点。
目前,节约饲料鱼粉用量的主要途径均是在寻找适宜的替代蛋白源及利用脂肪或糖类对蛋白的节约作用等基础上进行思考与探索,如在翘嘴鲌(Culteralburnus)[2]、齐口裂腹鱼(Schizothoraxprenanti)[3]和鲤(Cyprinuscarpio)[4-6]等均有这些方面工作的报道。开发植物蛋白源(如豆粕[2-3]、桑叶发酵蛋白[7]、玉米蛋白[8]等优质植物蛋白源)替代鱼粉的研究则是此类研究中较为活跃的部分,而其中集蛋白含量较高、数量体积庞大且经济实惠等优势于一身的豆粕则被认为是代替鱼粉的第一选择。但由于豆粕其本身含有例如胰蛋白酶抑制因子、大豆凝集素等抗营养因子,影响了动物肠道的微生态环境,营养物质的消化和吸收受到妨碍,再加上适口性较差,致使豆粕的使用在水产动物饲料中受到限制[9]。据报道,饲料中大豆分离蛋白替代40%的鱼粉时,虹鳟的生长及饲料转化率达到最佳[10];豆粕替代16%的鱼粉时牙鲆生长性能为适佳[11]。因此,寻找到在特定渔用饲料中豆粕替代鱼粉的适宜比例则成为此类研究的重点。目前,尚未见到有关植物蛋白质原料替代鱼粉在点带石斑鱼实用饲料中效果的研究报道。因此,本试验通过研究豆粕替代鱼粉对点带石斑鱼生长性能、消化能力及抗氧化能力的影响,评价豆粕替代鱼粉在点带石斑鱼实用饲料中的效果,旨在为优化点带石斑鱼人工配合饲料及合理利用大豆蛋白源提供依据。
1 材料与方法
1.1 试验饲料
配制6种蛋白质含量为48%、脂肪含量为12%、能量为19 MJ/kg的等氮等能饲料,对照组D0含60%蛋白质的鱼粉,试验组D1~D5组分别用豆粕代替10%、20%、30%、40%、50%的鱼粉蛋白,试验原料预混料和鱼油除外,用电动粉碎机粉碎,过60目网筛,经购自章丘市华祥颗粒机械有限公司的“华祥”牌SKJ-120型平模饲料颗粒机制成直径为2 mm的颗粒饲料,室温下风干,装入自封袋,-20 ℃保存备用,饲料配方和化学组成见表1。
1.2 试验鱼及饲养管理
试验鱼为天津市海发珍品实业发展有限公司提供的同一批鱼种,试验前先用基础饲料驯化7 d,后选取体质健壮、规格一致[平均体质量(84±2.5)g/尾]的点带石斑鱼,放入蓝色养殖箱(80 cm×40 cm×42 cm)中,每箱放养40尾。饲养试验进行10周,试验期间每天饱食投喂2次(09:00和15:30),每天早上清除箱内粪便,并换水,每天换水量为总水量的 1/3。养殖试验期间水体温度为(26±1) ℃,溶解氧浓度为 8.0~10.0 mg/L,pH值为7.5~7.7。

表1 鱼饲料组成和营养水平(干物质基础)
注:a.1 kg预混料组成如下:复合维生素:维生素B121.7 g、维生素B26.8 g、泛酸钙 16.9 g、烟酰胺 25.4 g、维生素B61.7 g、叶酸 0.5 g、肌醇67.7 g、包膜维生素C 3.4 g、氯化胆碱169.3 g、维生素K 1.4 g、维生素E 3.4 g、维生素A 0.7 g、生物素0.17 g。复合矿物质:乳酸钙 221.5 g、K2PO4161.9 g、CaHPO4·2H2O 91.4 g、MgSO4·7H2O 89.4g、Na2HPO4·2H2O 59.1g、柠檬酸铁 20.1 g、ZnSO4·7H2O 2 g、CoCl2·6H2O 0.7 g、MnSO4·H2O 0.5 g、KI 0.1 g、CuSO4·5H2O 54.2 g、Na2O3Se 0.03g。b.1 kg混合氨基酸组成如下:异亮氨酸 82.3g、亮氨酸 0.25 g、赖氨酸 0.5 g、苯丙氨酸 0.04 g、苏氨酸 132.9 g、缬氨酸 126.6 g、蛋氨酸 158.2 g。
1.3 样本采集
养殖试验结束时,试验鱼禁食24 h,从每个养殖箱中随机取15尾鱼,测定鱼体质量、体长和全长,采取尾部静脉抽血方法,每2尾鱼的血液为1个样本,注入2 mL的离心管中,在 4 ℃ 下3 500 r/min离心30 min,取上层血清备用,后于冰盘上解剖,取肝胰脏、前肠、中肠及后肠,迅速转入-80 ℃低温冰箱内保存,以用于抗氧化酶活力及消化酶活力的测定。
1.3 试验方法
1.3.1 生长指标计算 于试验结束后饥饿24 h称量鱼体质量计算增质量率、饲料效率、成活率等:
体增质量(WG)=(末均质量-初均质量)/初均质量×100%;特定生长率(SGR)=[ln(末均质量)-ln(初均质量)]/天数×100%;
饲料转化效率(FCR)=饲料干质量/增质量×100%;
蛋白质效率(PER)=增质量/蛋白质投喂量×100%;
肥满度(CF)=体质量/体长×100%。
1.3.2 粗酶液的制备 肝胰脏、前肠、中肠及后肠在冰生理盐水中漂洗,除去血液,用滤纸拭干,用移液管移入9倍于组织块质量的预冷匀浆介质(0.85%生理盐水),在匀浆器中进行匀浆,在4 ℃下4 500 r/min离心15 min,取上层清液备用。
1.3.3 生化指标检测 消化酶活力检测均采用商业试剂盒(南京建成生物工程研究所),每个样品每种酶活力均重复测定3次;蛋白酶活力采用福林酚法测定;脂肪酶活力的测定采用比浊法;淀粉酶采用碘——淀粉比色法进行测定。采用商业试剂盒(南京建成生物工程研究所)对血清及组织中还原型谷胱甘肽(GSH)和丙二醛(MDA)含量、超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH-PX)和过氧化氢酶(CAT)活力及总抗氧化能力(T-AOC)进行测定。其中GSH含量、SOD活力及T-AOC采用Thermofisher-4MK2型微孔读板机测定,其余指标采用UVmini-1240型紫外分光光度计测定。
1.4 数据统计
所有数据均用Excel 2007及SPSS 19进行分析处理。利用Excel软件制作各指标的标准曲线,并利用标准曲线计算得出各样品中对应指标的活性含量。所有数据均以“平均值±标准差”表示,并利用SPSS 19.0统计软件进行单因素方差分析(one-way ANOVA),若差异达到显著水平(P<0.05),则进行Duncan’s法多重比较。
2 结果与分析
2.1 豆粕替代鱼粉对点带石斑鱼生长性能的影响
由表2可知,随着替代水平的提高,平均末体质量、增质量率、特定生长率、蛋白质效率呈现明显的先上升后下降的趋势(P<0.05),其中,平均末体质量在10%~20%替代组中差异不显著,但均显著高于其他处理组;20%替代组的增质量率、特定生长率、蛋白质效率均显著高于其他处理组(P<0.05),其余处理组之间差异不显著。饲料系数随着替代水平的提高呈明显先下降后上升的趋势(P<0.05),其中饲料系数在10%~20%替代组中差异不显著(P>0.05),但显著低于其他处理组(P<0.05)。肥满度随替代水平提高显著上升(P<0.05),其中50%替代组与30%、40%替代组无显著差异,但显著高于其他处理组(P<0.05)。

表2 饲料中豆粕替代鱼粉比例对点带石斑鱼生长性能的影响
注:同列数据后不同小写字母表示显示显著差异(P<0.05)。下表同。
2.2 豆粕替代鱼粉对石斑鱼肠道消化酶活性影响
2.2.1 豆粕替代鱼粉对石斑鱼肠道蛋白酶活性的影响 由表3可知,前肠蛋白酶活性随着替代水平提高,先下降后升高,其中对照组D0(0%)与10%、40%替代组无显著差异,但显著高于其他对照组(P<0.05)。在中肠和后肠中蛋白酶活性随豆粕替代水平提高先升高后降低,其中中肠蛋白酶活性在40%替代组中显著高于对照组和10%替代组(P<0.05),但与其他处理组无显著差异;在后肠中40%替代组蛋白酶活性显著高于对照组和30%替代组(P<0.05),但与其他对照组无显著差异(P>0.05)。

表3 豆粕替代鱼粉对点带石斑鱼肠道蛋白酶活性
2.2.2 豆粕替代鱼粉对石斑鱼肠道淀粉酶活性的影响 由表4可知,投喂不同豆粕替代水平的饲料后,各组织中淀粉酶活性均值有一定波动。在前肠中20%组淀粉酶活性最高,显著高于对照组和30%替代组(P<0.05),与其他各组之间差异不显著。在中肠中40%与50%替代组无显著差异,但显著低于其他处理组(P<0.05)。在后肠中50%替代组淀粉酶活性与10%替代组无显著差异,但明显高于其他处理组。
2.2.3 豆粕替代鱼粉对石斑鱼肠道脂肪酶活性的影响 由表5可知,投喂不同豆粕替代水平饲料后,肠道中脂肪酶活性先升高后降低。在前肠、中肠、后肠中20%替代组明显高于其他处理组(P<0.05)。
2.3 豆粕替代鱼粉对石斑鱼抗氧化指标影响
由表6可知,随着替代水平的提高,血清超氧化物歧化酶(SOD) 活性显著降低, 最高值出现在对照组, 显著高于50%替代组(P<0.05),与其他处理组之间差异不显著;在肝脏中D2(20%)组SOD活性最高且与其他5组存在显著差异(P<0.05)。过氧化氢酶活性随豆粕替代量增加先增强后减弱,肝脏中过氧化氢酶活性最高为D2组且与其余5组存在显著差异(P<0.05);而血清中过氧化氢酶活性最高为D4组,与其余5组差异不显著(P>0.05)。在血清中,丙二醛含量随豆粕替代量的变化并无明显变化,豆粕替代鱼粉对丙二醛影响并不显著(P>0.05);在肝脏中,D2组丙二醛含量最高,且与其他几组差异明显(P<0.05)。还原型谷胱甘肽活性均值有一定的波动,在血清中D1组GSH表达量最高但与其他5组无显著差异(P>0.05);在肝脏中D4组GSH含量最高,与其余5组存在显著差异(P<0.05)。总抗氧化能力均值有一定的波动,在血清中 T-AOC最高组为D3但与其余5组无显著差异(P>0.05);T-AOC在肝脏中最高位D2组,与其他5组存在明显差异(P<0.05)。

表4 豆粕替代鱼粉对点带石斑鱼肠道淀粉酶活性

表5 豆粕替代鱼粉对点带石斑鱼肠道脂肪酶活性
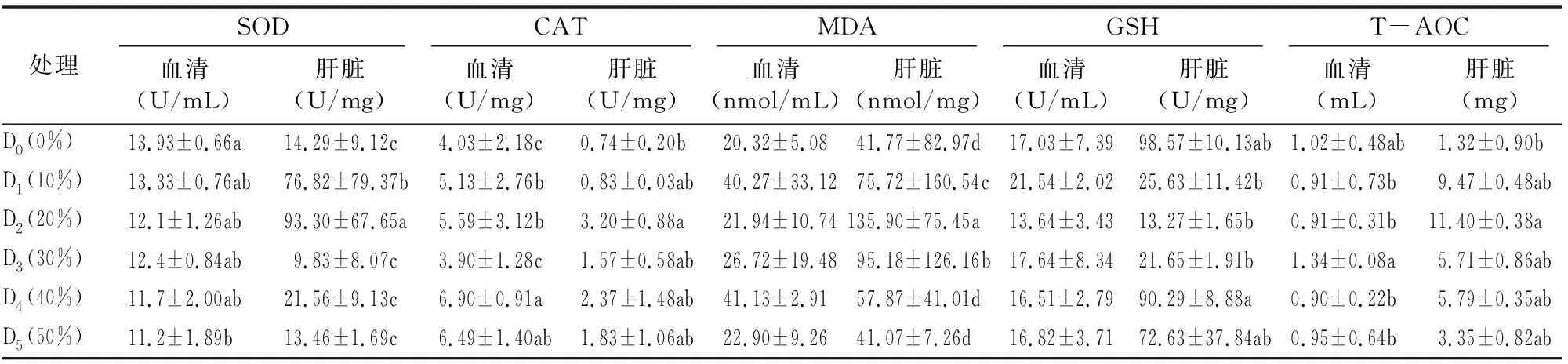
表6 豆粕替代鱼粉对石斑鱼抗氧化指标影响
3 讨论
3.1 饲料中豆粕替代鱼粉比例对点带石斑鱼生长性能的影响
蛋白质是鱼类生长过程中的关键因素,是构成动物机体的成分,同时还为水产动物提供能量。目前,在水产动物饲料中植物蛋白源部分替代鱼粉成为研究的热点。已有相关研究报道,当膨化豆粕替代鱼粉蛋白比例为60%时,不会对虹鳟鱼生长指标产生影响[12]。在5%豆粕替代鱼粉用量的1/3时,不影响奥尼罗非鱼幼鱼及鱼种生长性能,但当替代量达2/3时,幼鱼生长显著降低[13]。本试验中,饲料中豆粕替代鱼粉对点带石斑鱼增质量率、蛋白质效率和特定生长率有显著影响,且随豆粕的替代量的增大呈先增后减的趋势,在豆粕替代水平为20%的D2组,增质量率、特定生长率和蛋白质效率均为最大值,且饵料系数降为最低。而当替代水平再上升时,增质量率、特定生长率和蛋白质效率都随之下降,而饵料系数却随之上升,这与邹文超等的研究结果[14]相符,其研究结果表明斜带石斑鱼饲料中豆粕替代鱼粉蛋白的比例不宜超过30%,当豆粕替代40%鱼粉蛋白时,斜带石斑鱼生长性能下降。从上述2个试验中可以发现,在石斑鱼饲料中豆粕替代鱼粉比例适宜时,不但不会影响其生长,反而会在一定程度上增强其饲料利用能力,进而促进生长;但替代比例过高,豆粕存在的多种抗营养因子、败风味物质可能造成饲料适口性不佳,使得饲料中较高的豆粕比例降低了鱼类对饲料蛋白质的消化能力,并且其氨基酸利用效率较低,尤其是蛋氨酸、赖氨酸利用率不足,影响水生动物对饲料的利用,抑制生长。而杨严鸥等指出在不影响黄颡鱼生长性能的前提下,其饲料中豆粕替代鱼粉蛋白含量最高为30%,但过量替代时,黄颡鱼的生长指标同样会降低,上述替代量的差异可能是由于试验鱼种类不同,养殖条件存在差异等原因[15]。
肥满度被视为鱼类生物学研究中衡量生长和摄食强度等健康状况的1个指标,其变化也预示着鱼体对于饲料营养的利用情况。本研究发现,肥满度随替代水平提高显著上升,而刘汝鹏等及王桂芹等对翘嘴鲌的研究发现豆粕替代鱼粉水平未对肥满度产生显著性影响,但亦成升高趋势[16-17]。综合上述生长指标分析发现,适宜豆粕替代鱼粉比例能够增强鱼的摄食强度,进而使鱼体获得更好的生长及肥满度;但替代比例过高,随肥满度仍呈升高趋势,但鱼体生长性能出现下降,表明从表观来看鱼体获得良好的肥满度,并不是因为饲料利用能力的增强,而可能是因为饲料蛋白质或糖转变为脂肪或者糖原蓄积在肝脏或内脏中等原因造成的,其具体的影响机制仍需进一步研究。
3.2 饲料中豆粕替代鱼粉比例对点带石斑鱼消化酶活性的影响
在豆粕替代鱼粉的过程中,胰蛋白酶抑制因子、皂角苷、大豆凝集素等豆粕自身含有的抗营养因子均会成为抑制水产动物蛋白酶分泌及活性的重要因素,如Sveier在对大西洋鲑鱼的研究中发现胰蛋白酶抑制因子既可以通过在肠道中与蛋白酶直接结合的方式降低蛋白酶活性,又可以通过影响肝胰脏发育抑制蛋白酶的合成与分泌[18]。在肠道中,前肠作为其中主要消化蛋白的部分,本研究中发现豆粕替代水平的升高会降低点带石斑鱼前肠蛋白酶活性,而中肠及后肠蛋白酶活性则随之呈显著上升趋势,但肠道蛋白酶活性总体仍呈下降趋势,推测鱼体会通过调节肠道中不同部位的蛋白酶合成及分泌来适应饲料中蛋白源的变化。这与邹文超对斜带石斑鱼(Epinepheluscoioides)的研究结果[19]相似。此外,本试验发现蛋白源变化在引起蛋白酶活性变化的同时,也会引起使其他消化酶活性产生相应变化,如当豆粕替代鱼粉蛋白比例达到20%时会显著提高前肠中淀粉酶活性,揭示豆粕替代比例的升高会增加饲料中的淀粉含量,进而诱导鱼体内淀粉酶的合成。而钱曦等在对翘嘴鲌(ErythroculterilishaeformisBleeker)的研究中发现,肠道淀粉酶活性并不因豆粕替代鱼粉比例的提高而有显著性变化[20],究其主要原因是鱼种的差异,相较于肉食性的点带石斑鱼,杂食性的翘嘴鲌体内的淀粉酶活性较高[21],能够更好地适应饲料中淀粉含量的增加。此外,当豆粕替代鱼粉比例达到20%时也会显著提高与前肠、中肠及后肠的脂肪酶活性,其具体影响机制还有待进一步研究。
3.3 饲料中豆粕替代鱼粉比例对点带石斑鱼抗氧化能力的影响
鱼类体内抗氧化酶的缺乏会导致体内原有的自由基代谢平衡遭到破坏;正常情况下,体内产生的自由基即活性氧可被非酶性或酶性抗氧化系统清除掉,其中SOD和GSH-Px是鱼体内的主要抗氧化酶,SOD能催化超氧阴离子产生歧化反应,清除超氧阴离子,保护细胞免受损伤。GSH-Px能特异地催化还原型谷胱甘肽(GSH)对H2O2的还原反应,保护细胞膜结构和功能的完整性[22-23]。在本试验的豆粕替代梯度中,各替代梯度的血清的抗氧化能力指标基本没有明显变化,而肝脏中GSH值先减后增,SOD与MDA值则是先增后减。综合来看,豆粕替代20%鱼粉对点带石斑鱼的抗氧化能力比较有利。上述研究表明,在点带石斑鱼饲料中用豆粕部分替代鱼粉,可改善鱼体抗氧化能力,且在适当替代范围内抗氧化能力的改善程度与替代比例间存在同步性,但替代量过高后豆粕自身含有的抗营养因子除了会影响鱼类消化能力外,也会引起机体的免疫反应或是炎症的产生,进而抑制鱼体非特异性免疫性能,这与对斜带石斑鱼(Epinepheluscoioides)[21]及中华鳖(TrionyxSinensis)[24]的研究结果一致。
4 结论
本研究中,以豆粕替代20%鱼粉蛋白时,能显著提高点带石斑鱼的生长性能又不会对点带石斑鱼的消化酶活性及抗氧化能力产生明显影响;而且从生长性能、消化酶和抗氧化能力不受影响的角度考虑,点带石斑鱼饲料中豆粕替代鱼粉蛋白的比例不宜超过20%。
