支链淀粉结构对稻米淀粉糊化特性的影响
2018-09-03周慧颖彭小松欧阳林娟朱昌兰陈小荣傅军如边建民胡丽芳贺浩华贺晓鹏
周慧颖 彭小松 欧阳林娟 朱昌兰 陈小荣 傅军如 边建民 胡丽芳 贺浩华 贺晓鹏
(江西农业大学农学院;作物生理生态与遗传育种教育部重点实验室; 双季稻现代化生产协同创新中心,南昌 330045)
直链淀粉含量(Amylose Content,AC)对稻米食味品质有重要影响[1-2],而支链淀粉精细结构的不同是导致AC相似品种间品质差异的重要因素[3-4]。支链淀粉精细结构包括链长和链长分布、平均链长(Average Chain Length,CL)、平均外链长(Average exterior Chain length,ECL)、平均内链长(Average Internal Chain Length,ICL)、A∶B值等参数[5]。在支链淀粉链长和链长分布与稻米品质和淀粉理化特性关系的研究方面,胡志萍等[6]研究表明,淀粉的链长及支链淀粉长、中、短链的比例不同,可以使相同AC的品种可能具有不同的糊化温度和胶稠度。贺晓鹏等[7]研究指出,支链淀粉不同链长范围的支链数量比例主要与淀粉的糊化温度相关,而与淀粉的胶稠度和RVA成糊特性关系不密切。还有报道,Vandeputte等[8]认为支链淀粉DP为6~9的短链引起糊化温度下降;DP为12~22的链使得起始糊化温度(Onset Gelatinization Temperature,To)上升;DP值为6~9和DP>25的支链淀粉链降低了支链淀粉终结糊化温度(End Gelatinization temperature,Te)及糊化焓(Gelatinization Enthalpy,ΔH)。除链长和链长分布外,稻米支链淀粉的CL、ECL、ICL、A∶B值等结构参数的研究相对较少。
本研究选取了26个AC为11%~15%的低AC类型水稻品种,应用改进的酶法测定了支链淀粉的β-淀粉酶水解率、CL、ECL、ICL、A∶B值等结构参数,探讨了支链淀粉的精细结构与稻米淀粉RVA成糊特性及DSC热特性的相关性,以明确支链淀粉结构对稻米品质的影响。
1 材料与方法
1.1 材料与仪器
选取26个AC为11%~15%的低AC类型水稻品种,其中籼稻品种8个,粳稻品种18个,具体见表1。各供试品种于2014年在江西农业大学实验田作中稻种植,均按常规水肥管理,分期播种,同期收获。水稻种子成熟后收获晒干,用精米机充分研磨成精米后进一步磨成米粉,通过100目的筛子,充分干燥后密封保存,用于后续研究。
β-淀粉酶(EC 3.2.1.2)、普鲁兰酶(EC 3.2.1.41)、异淀粉酶(EC 3.2.1.68):Sigma-Aldrich公司。
SCINO-CT410旋风式样品磨:瑞典FOSS公司;TU-1810D紫外可见分光光度计:北京普析公司;RVA-TecMaster黏度速测仪:瑞典Perten公司;DSC4000差示扫描量热仪:美国Perkin-Elmer公司。
1.2 支链淀粉精细结构的测定
利用基于β-淀粉酶、异淀粉酶、普鲁兰酶的酶法并加以优化后进行支链淀粉CL、ECL、ICL、A∶B值的测定[9]。
1.2.1 支链淀粉的分离与纯化
支链淀粉的纯化参照贺晓鹏等[7]的方法。
1.2.2 β-淀粉酶水解率的测定
称取0.5 g纯支链淀粉放入50 mL的离心管中,加入浓度为0.02 mol/L,pH 5.0的醋酸缓冲液25 mL,充分混匀后加入β-淀粉酶2 500 U,同时滴加1~2滴甲苯防止微生物生长,然后37 ℃水浴保温反应时间48 h后,将反应液置于沸水浴中25 min灭酶,冷却后测定体系中的总糖和总还原力,以不加酶的样品作为对照,计算支链淀粉β-淀粉酶水解率:
β-淀粉酶水解率=还原力(麦芽糖当量)×100/总糖(麦芽糖当量)
1.2.3 支链淀粉链长的测定
称取22 mg纯支链淀粉放入5 mL离心管中,加入浓度为0.05 mol/L,pH 5.0的醋酸缓冲液5 mL,充分混匀后加入普鲁兰酶4 U,液面以1~2滴甲苯覆盖后置于37 ℃的水浴中保温。反应24 h后,将反应液沸水浴灭酶20 min,冷却后利用蒽酮-硫酸法和二硝基水杨酸(DNS)法测定体系中的总糖和还原糖含量(μg/mL),以不加酶的样品作为对照计算CL,并计算出ECL和ICL:
CL=产物中的总糖(葡萄糖当量)/产物中的总还原力(葡萄糖当量)
ECL=CL ×β-淀粉酶水解率+2.0
ICL=(CL-ECL)-1.0
1.2.4 支链淀粉A∶B值的测定
将测定β-淀粉水解率时的最终反应液加入3倍体积甲醇,冰浴30 min后6 000 r/min离心5 min,倾去上清液,加入75%的甲醇冲洗沉淀,37 ℃恒温干燥后用研钵磨成细粉,即得支链淀粉β-极限糊精。称取5 mg支链淀粉β-极限糊精于2 mL离心管中,加入200 μL蒸馏水,沸水浴中加热搅拌10 min。再次加入浓度为0.04 mol/mL,pH4.5的醋酸缓冲液,沸水浴中加热搅拌10 min后冷却至室温,制成质量浓度为1 mg/mL的β-极限糊精溶液。以此为底物,取2 mL加入异淀粉酶100 U/mL,在37 ℃条件下振荡反应24 h后,置沸水浴中加热20 min灭活异淀粉酶,利用DNS法测定其还原糖含量C1(μg/mL)。继续往反应液中加入普鲁兰酶1 U/mL,在37 ℃条件下振荡反应24 h后,置沸水浴中加热20 min灭活普鲁兰酶,测定其还原糖含量C2(μg/mL),即可计算出A∶B值:
C1=B+0.5A
C2=B+A
1.3 稻米RVA成糊特性的测定
RVA成糊特性采用黏度速测仪测定,按AACC(美国谷物化学家协会)操作规程,并用TCW配套软件进行数据分析[10]。称取实验样品3.0 g放入铝罐内,加入25 mL蒸馏水,充分混匀后,放入仪器中进行糊化。具体测定过程如下:先在50 ℃下保持1 min,然后在3.75 min内恒速升温至95 ℃,并保持2.5 min,再以恒速在3.75 min内降温到50 ℃,并且保持1.4 min。搅拌器在起始10 s内转动速度为960 r/min,之后保持在160 r/min,从测定开始到结束共需12.5 min。RVA成糊特性曲线用最高黏度(PKV)、热浆黏度(HPV)、冷胶黏度(CPV)、崩解值(BDV,PKV-HPV)、消减值(SBV,CPV-PKV)等特征值来表示,每个样品重复测定3次。
1.4 DSC热特性的测定
DSC热特性用差示扫描量热仪测定。称取5.0 mg左右实验样品置于铝盒中,并记录其具体重量,然后加入15 μL的蒸馏水,用配套铝盖密封铝盘,在室温下平衡过夜。以只加蒸馏水的铝盘作为参比,测定处理样品,以10 ℃/min的升温速率从30 ℃上升到110 ℃,得到试样的DSC热特性曲线。To、最高糊化温度(Peak gelatinization temperature,Tp)、Te及糊化时的ΔH等参数在配套通用分析软件中分析得到[11],每个实验样品重复3次。
1.5 数据分析
采用SPSS17.0统计软件对实验数据进行统计分析[12],对全部品种和AC为11%~13%及AC为13%~15%两组的支链淀粉结构与RVA成糊特性及DSC热特性的相关性进行分析。
2 结果与分析
2.1 不同水稻品种支链淀粉结构和淀粉糊化特性
26个参试品种的支链淀粉结构参数和RVA成糊特性及DSC热特性参数的测定结果见表1和表2。进一步分析参试水稻品种的支链淀粉精细结构参数之间的相关性(表3)发现,在支链淀粉结构参数之间除了ICL与CL、A∶B值间无显著的相关性间,其他参数两两之间均检测到了显著的相关性,可见支链淀粉不同结构参数之间相互都有关联。
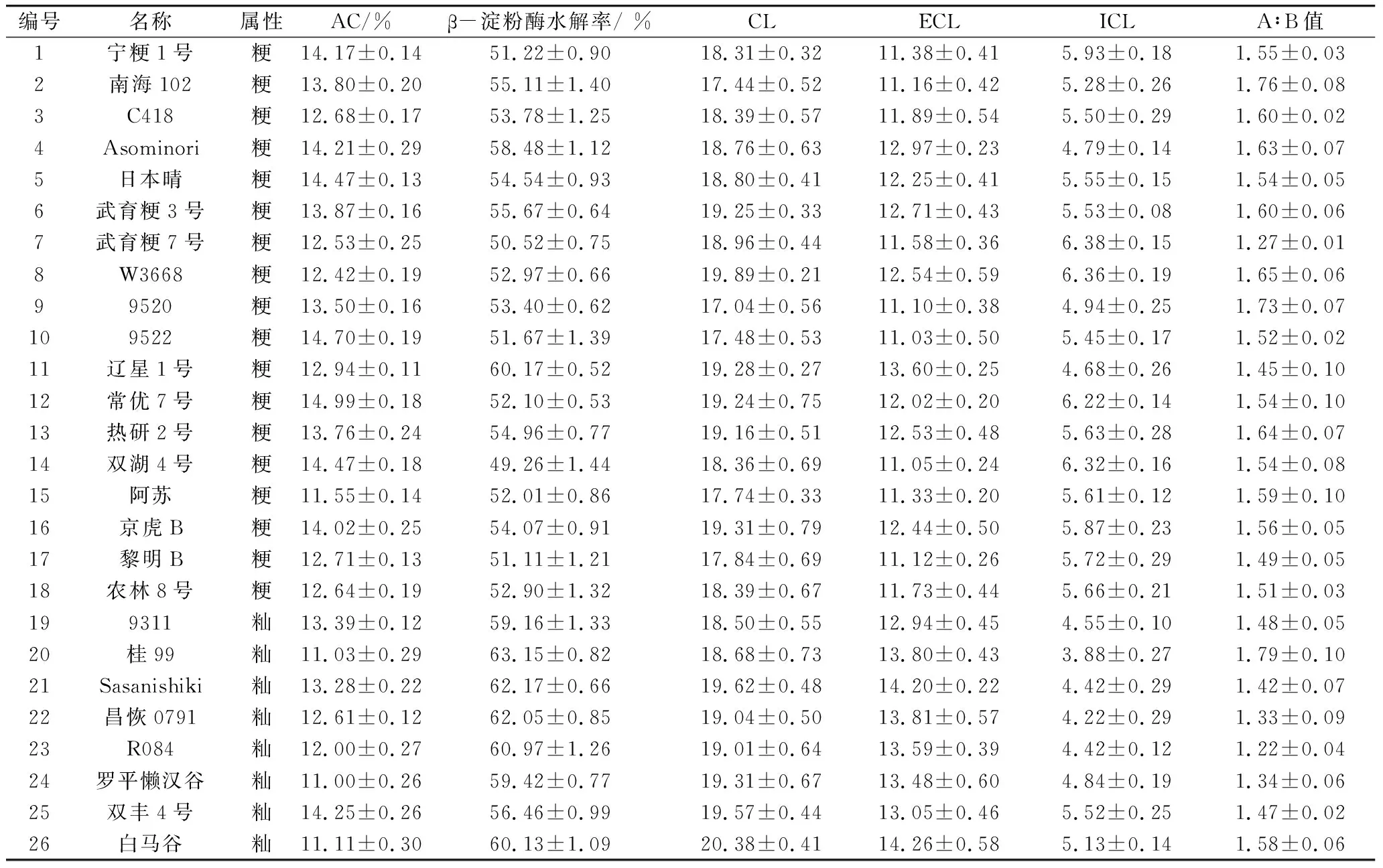
表1 26份低AC水稻品种AC及其结构参数

表3 支链淀粉结构间的相关性
注:*、 ** 分别为达显著(P<0.05)和极显著水平(P<0.01),下同。
2.2 支链淀粉结构与淀粉糊化特性参数的相关性
支链淀粉精细结构和淀粉RVA成糊特性及DSC热特性间的相关性见表4,由表4可知,26个水稻品种中ECL与DSC热特性中的To、Tp呈极显著正相关,与Te也呈显著正相关;与RVA成糊特性中的PKV呈极显著负相关,与SBV呈极显著正相关,与HPV、BDV呈显著负相关。CL与To、Tp、Te呈显著正相关;与PKV、BDV呈极显著负相关,与SBV呈极显著正相关。A∶B值与To呈显著负相关,与PKV、BDV呈显著正相关,与SBV呈显著负相关。ICL只与To呈显著负相关,与其他参数间的相关性不明显。为尽量排除AC对稻米淀粉糊化特性的影响,将参试的26个品种按AC分成11%~13%(12个品种)和13%~15%(14个品种)两组,由表4可知,两组材料的支链淀粉结构与淀粉RVA成糊特性及DSC热特性的相关性与全部26个品种间的基本一致。具体表现为,ECL在两组材料中与To、Tp呈显著正相关;与PKV呈显著负相关,与SBV呈显著正相关。CL在两组材料中与Tp呈显著正相关;与PKV、BDV在11%~13%分组和13%~15%分组间分别呈极显著和显著负相关;与SBV在11%~13%分组和13%~15%分组间分别呈极显著和显著正相关。A∶B值与To呈显著负相关,与PKV、BDV呈显著正相关,与SBV呈显著负相关。
3 讨论
支链淀粉作为稻米淀粉的最主要组分,是淀粉粒各级结构形成和稻米理化特性的主要决定因素,也是造成AC相同或相近的水稻品种间稻米品质差异的主要原因。现有研究表明,稻米淀粉的糊化过程,实质上就是淀粉晶体的溶解,其中涉及了淀粉体形状的差异、淀粉体间排列的紧密程度、直链淀粉从淀粉体中的浸出、支链淀粉链的分子组成、淀粉晶体中支链淀粉双螺旋结构的断裂和分解等[13]。在本研究中,26个水稻品种的CL、ECL与To、Tp、Te呈正相关,A∶B值与To呈负相关,这可能是由于稻米支链淀粉的短链比例高时,由于短链不能最大程度地堆积到晶体片层中去,晶体化顺序较差,因而To、Tp、Te低;而当支链淀粉中的长链比例高时,这些长链可能通过形成较长的双螺旋结构,从而导致糊化时需要较高的温度来解离,因而To、Tp、Te高[14-15]。同时,相对较高To、Tp、Te的稻米品种具有相对紧密的组织结构,在淀粉糊化时水分渗透、扩散到淀粉颗粒内部的能力和速度较慢,因此支链淀粉的长链所占比率越高的稻米品种在蒸煮时需要更多的水分和更长的蒸煮时间[16-17]。
近年来,RVA成糊特性与稻米品质的关系已有了一定的研究[18-21],在本研究中,26个水稻品种的CL、ECL与PKV和BDV呈负相关,A∶B值与PKV和BDV呈正相关,这可能是由于支链淀粉中长链的比率越高,分子间的作用力就越强,抑制了淀粉的膨胀,蒸煮性变差,不利于淀粉的糊化,PKV降低;同时长链也使糊化过程中支链淀粉结构不易破裂,使淀粉颗粒结构维持在胶稠化状态,抑制了淀粉糊液黏度的下降,从而降低了BDV,米饭质地变硬,食味品质下降[22-23]。研究还发现,CL、ECL与SBV呈正相关,而A∶B值与其呈负相关,这是由于淀粉糊化过程后期,随着温度的降低,米糊中的直链淀粉和支链淀粉再度进行分子间的重排,因此,支链淀粉中长含量越高,淀粉中的分子重新排列越容易形成恢复成有序状态,SBV增大;而支链淀粉中短链含量越高,其分子链间的松散程度越高,从而减弱了淀粉分子链重新缔合的紧密程度,SBV减小,米饭冷却后表现出良好的抗回生能力。
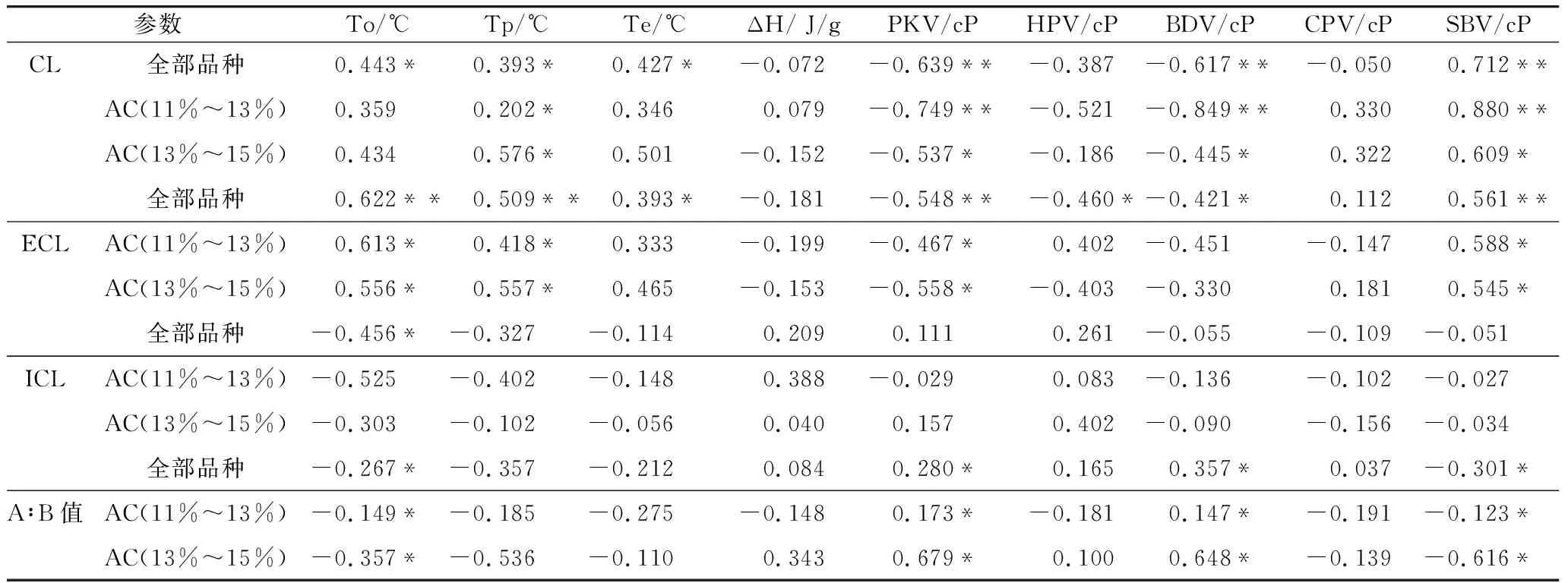
表4 支链淀粉结构与淀粉糊化及热特性参数间的相关性
4 结论
支链淀粉结构的不同是导致AC相近的水稻品种间品质差异的主要原因,淀粉的链长结构是影响淀粉糊化的主要因素,支链淀粉的CL、ECL、A∶B值对淀粉RVA成糊特性及DSC热特性有较大影响。26个水稻品种的CL、ECL与To、Tp、Te、SBV呈正相关,与PKV和BDV呈负相关;A∶B值与PKV和BDV呈正相关,与To、SBV呈负相关,因此,低CL、ECL及高A∶B值的稻米淀粉RVA成糊特性较好、糊化温度较低。
