发酵豆粕替代鱼粉对黄姑鱼幼鱼肌肉氨基酸,IGF-I基因相对表达量及肝脏组织结构的影响
2018-08-20胡沈玉王立改曾文繁陈睿毅谢庆平徐冬冬
胡沈玉,王立改,楼 宝,曾文繁,谭 朋,詹 炜,陈睿毅,刘 峰,谢庆平,徐冬冬
(浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江舟山 316021)
黄姑鱼Nibea albiflora属鲈形目石首鱼科黄姑鱼属,俗称黄婆鸡、黄姑子、铜锣鱼等,其生长快,抗病力强,营养丰富,是我国重要的海洋经济鱼类。由于环境遭到破坏和过度捕捞,黄姑鱼的自然资源不断减少[1-2]。前人相继在黄姑鱼的早期生活史[3]、黄姑鱼胚胎及仔稚鱼形态特征[4]、人工育苗养殖[5-6]等方面进行了研究。而在黄姑鱼饲料营养方面,一些学者对黄姑鱼饲料中维生素[7]、脂肪[8-9]、蛋白质水平[10-11]需求等方面进行了研究,目前,发酵豆粕替代鱼粉的研究尚未见报道。鱼粉是黄姑鱼饲料中的重要组成成分,近年来鱼粉供应不足导致其价格不断上涨,提高了黄姑鱼养殖成本。因此寻找质优价廉的蛋白源成为当务之急。发酵豆粕是利用有益微生物发酵豆粕后获得,有效消除了抗营养因子,降低了大、中分子蛋白水平,提高了小分子蛋白含量及氨基酸总量,大大改善了豆粕的营养价值[12]。本试验以黄姑鱼为研究对象,在黄姑鱼配合饲料中以不同比例的发酵豆粕替代鱼粉,研究其对黄姑鱼幼鱼肝脏和肌肉中IGF-I基因表达量及肌肉中氨基酸含量的影响,为黄姑鱼饲料中用发酵豆粕替代鱼粉提供科学依据。
1 材料和方法
1.1.实验设计和样品采集
试验以发酵豆粕、豆粕、鱼粉和小麦蛋白粉为蛋白质源,鱼油、豆油和大豆卵磷脂为主要脂肪源,配制含45%鱼粉的基础饲料。以发酵豆粕替代基础饲料中0%(FSM0组)、10%(FSM10组)、20%(FSM20组)、30%(FSM30组)、40%(FSM40组)、50%(FSM50组)的鱼粉,共配制6组饲料。试验饲料配方见表1,饲料氨基酸成分见表2。原料经粉碎后过60目筛,按配方称重,采用逐级扩大法使之混合均匀,放于搅拌机中,搅拌的同时加入适量水分,原料搅拌均匀后放入F-26型双螺杆挤条机制成粒径为2 mm和4 mm的配合饲料,自然风干到水分含量大约为10%,分装后存于-20℃冰箱备用。
实验用的黄姑鱼取自西轩渔业科技岛试验场人工培育苗种。初步选择600余尾同龄黄姑鱼幼鱼暂养于室内50 m3水泥池内,暂养2周后,选取健康活跃、大小均匀、平均体重为(31.24±0.02)g的360尾鱼随机分成6组,每组设置3个重复,每个重复20尾,饲养在500L水桶中。将6种饲料和18桶鱼随机分配,每天07:00和16:00各投喂1次。喂食量为鱼体重的3%~5%,并每过2周称重1次以调整喂食量,饲养8周。饲养期间,水温(27±2)℃,盐度28~29。实验结束后每桶随机选取3尾鱼,取肌肉、肝共2种组织样品置于RNA保存液中,用于IGF-I基因表达量的测定;取足够量的肌肉用于氨基酸组成的分析,于-80℃保存备用;并取FSM0,FSM20和FSM50组的肝脏固定于波恩氏液中,用于肝脏组织学观察。
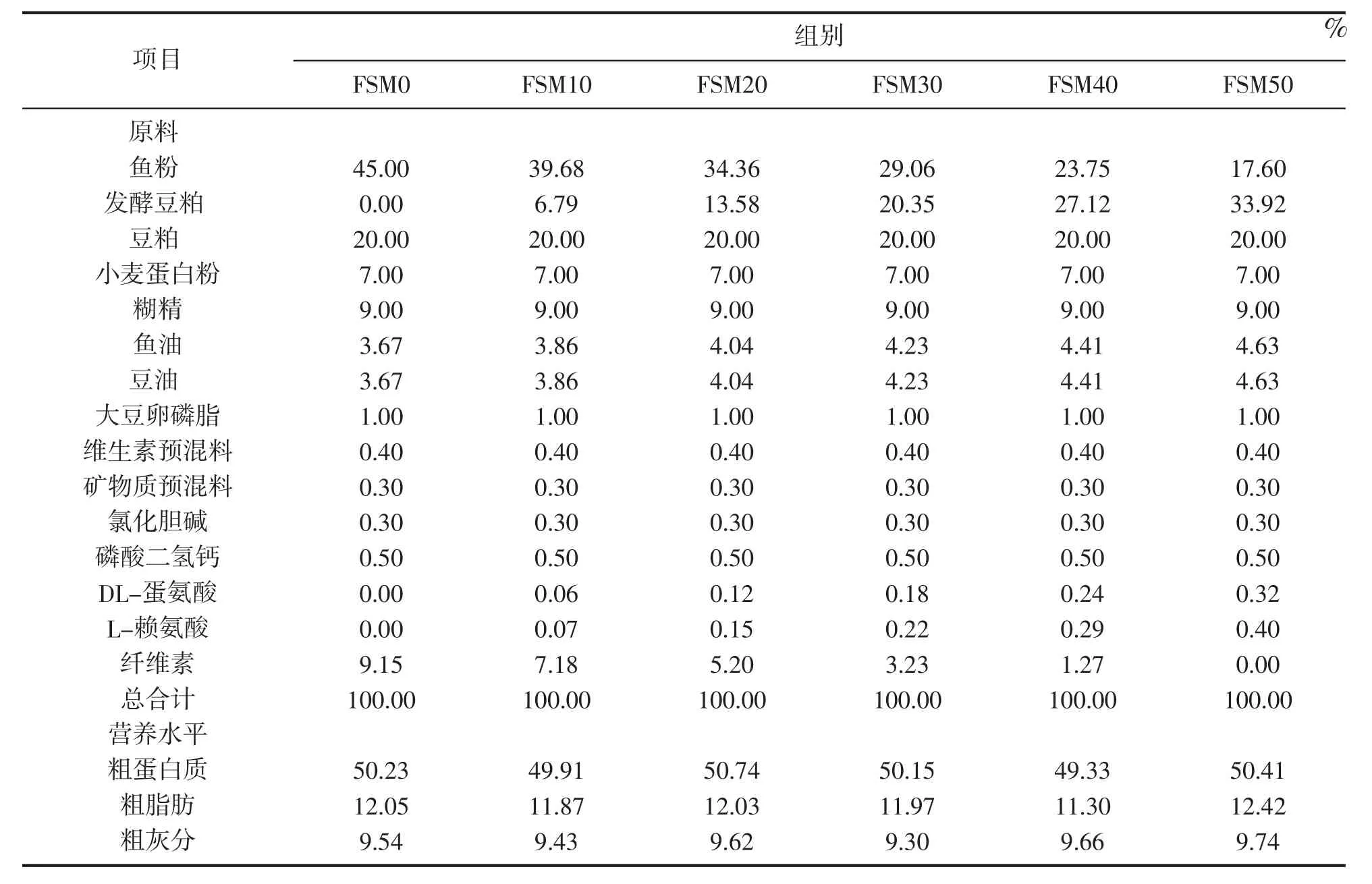
表1 试验饲料组成及营养水平(干物质基础)Tab.1 Composition and nutrient levels of experimental diets(DM basis)
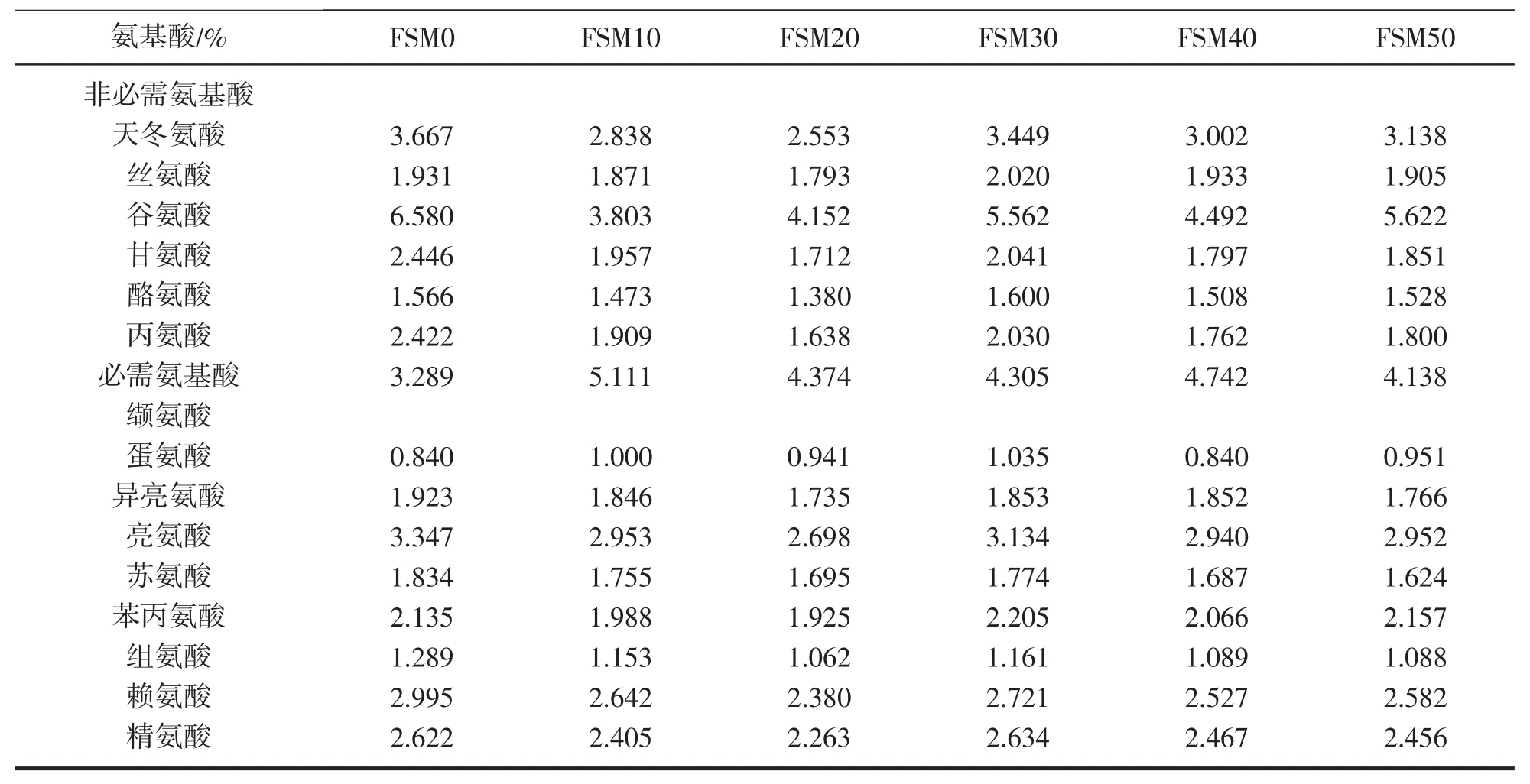
表2 饲料的氨基酸含量分析(饲料干物质%)Tab.2 The amino acid contents in the diets(Dry matter%)
1.2 总RNA提取及反转录
取组织样品50 mg于液氮中研磨,加入1 mL裂解液,按(Solarbio)总RNA提取试剂盒说明书提取总RNA。在紫外分光光度计下测定其纯度和浓度,OD260/280的值在1.8~2.0之间,说明提取的总RNA质量较好。对提取总RNA做琼脂糖凝胶电泳,1%的胶浓度,110 V,15 min,以此来检查总RNA完整性。使用全式金逆转录试剂盒进行逆转录。
1.3 实时荧光定量PCR反应
以大黄鱼的β-actin基因为持家基因,根据前期克隆得到黄姑鱼IGF-I全长基因序列(MG004542)设计荧光定量PCR引物,引物由上海生工生物工程有限公司合成,引物序列见表3。

图1 RNA电泳图Fig.1 RNA electrophoresis

表3 荧光定量PCR引物Tab.3 Real-time PCR primers
1.4 qRT-PCR反应的条件
qRT-PCR选用SYBR Green染色法,采用TransStart®Tip Green qPCR SuperMix试剂盒在ABI StepOnePlus实时荧光PCR系统下进行,采用三步法以20 uL体系进行qRT-PCR,在94℃下预变性30 s,94℃变性5 s,59℃退火30 s,72℃延伸10 s,循环数为40。反应结束后在60~95℃插入溶解曲线。
1.5 肌肉氨基酸含量测定
用HP1100高效液相色谱法(HPLC 1100,CA,美国)测定氨基酸含量。
1.6 肝脏组织形态学测定
取用70%乙醇保存的肝脏,在酒精中梯度脱水后用石蜡包埋,切片后用苏木精-伊红(HE)染色法染色,后用显微镜观察肝脏组织切片的结构。
1.7 数据统计分析
通过2-ΔΔCt法计算黄姑鱼IGF-I表达量,并使用SPSS19.0软件对结果进行单因子方差分析(one-way ANOVA)。同时对数据进行多重比较检验,以分析不同发酵豆粕配比下IGF-I表达量和氨基酸水平差异性当P<0.05为差异显著。数据用平均值±标准差(Mean±SD)的形式来表示。
2 结果
2.1 不同水平的发酵豆粕替代鱼粉对黄姑鱼幼鱼肌肉氨基酸组成的影响
各组肌肉中的氨基酸组成中(表4),缬氨酸和蛋氨酸均无显著性差异,其余氨基酸均随着发酵豆粕替代水平的提高而呈现增高的趋势。其中FSM30、FSM40和FSM50这3组的肌肉氨基酸组成显著高于FSM0组(P<0.05)。
2.2 不同水平的发酵豆粕替代鱼粉下黄姑鱼幼鱼肝脏IGF-I基因的相对表达量
肝脏中IGF-I的相对表达量采用2-ΔΔCt法计算,FSM0组的表达量设定为1,如图2。从图2可以看出,FSM0组IGF-I基因表达量最高,FSM50组IGF-I基因表达量显著低于对照组。

图2 发酵豆粕替代不同比例鱼粉对黄姑鱼幼鱼肝脏中IGF-I基因相对表达量的影响Fig.2 The effects of replacing fish meal with fermented soybean meal on liver IGF-I expression of juvenile N.albiflora

表4 不同水平的发酵豆粕替代鱼粉对黄姑鱼幼鱼肌肉氨基酸组成的影响(%)(平均值±标准差)Tab.4 The effects of replacing fish meal with fermented soybean meal on amino acids in muscle(Mean±SD)
2.3 不同水平的发酵豆粕替代鱼粉下黄姑鱼幼鱼肌肉IGF-I基因的相对表达量
肌肉中IGF-I的相对表达量采用2-ΔΔCt法计算,FSM0组的表达量设定为1,如图3。肌肉中IGF-I基因的表达量呈先上升后下降的趋势。其中FSM30表达量显著高于其它各组。
2.4 不同水平发酵豆粕替代鱼粉对黄姑鱼幼鱼肝脏组织形态的影响
肝脏组织观察,如图4所示,随着发酵豆粕替代水平的增加,肝细胞空泡化加剧,肝细胞内脂肪增多。FSM0组细胞排列规则整齐,细胞核正常;FSM20组的肝细胞出现少量空泡化;而FSM50组肝脏细胞空泡化现象严重,肝细胞轮廓模糊,部分肝细胞核已经消失不见。

图3 发酵豆粕替代不同比例鱼粉对黄姑鱼幼鱼肌肉中IGF-I基因相对表达量的影响Fig.3 The effects of replacing fish meal with fermented soybean meal on muscle IGF-I expression of juvenile N.albiflora
3 讨论
3.1 不同水平的发酵豆粕替代鱼粉对黄姑鱼幼鱼肌肉氨基酸组成的影响
豆粕经微生物发酵可有效降低抗营养因子含量,将大分子蛋白质降解为小肽和游离氨基酸,钝化抗营养因子活性,提高饲料转化效率[13-15]。崔燕燕等[16]表明中华绒螯蟹Eriocheir sinensis饲料在发酵豆粕替代50%的鱼粉后,发酵豆粕组总必需氨基酸的沉积率比鱼粉组高。许文婕等[17]发现异育银鲫Carassius auratus gibelio对发酵豆粕的总氨基酸表观消化率在12种饲料原料中是最高的。本实验黄姑鱼肌肉氨基酸分析结果显示(表2),在15种氨基酸中,除缬氨酸和蛋氨酸含量没显著性差异外,其它氨基酸含量都有差异,且随着发酵豆粕替代量的升高呈上升趋势。表明饲料中发酵豆粕部分替代鱼粉可以促进肌肉中大部分氨基酸的吸收积累,提高黄姑鱼肌肉中大部分氨基酸的沉积率。

图4 不同水平发酵豆粕替代鱼粉对黄姑鱼幼鱼肝脏组织形态的影响Fig.4 The effects of replacing fish meal with fermented soybean meal on liver tissue morphology of juvenile N.albiflora(HE staining,400×)
3.2 不同水平的发酵豆粕替代鱼粉对黄姑鱼幼鱼肝脏和肌肉IGF-I基因的相对表达量的影响
IGF-I对动物新陈代谢、生长繁殖和维持神经系统正常运作有十分重要的作用[18],其在动物体内各组织中均有表达[19-20],其中肝脏中表达量最高[21-23]。研究表明营养状态可以影响IGF的合成和分泌[24],禁食可降低肝脏IGF-I的基因表达量,而重新给鱼投喂食物后IGF-I基因表达量又随之上升[25]。本实验中,FSM50组肝脏中IGF-I表达量显著低于对照组(FSM0组),原因可能是随着发酵豆粕替代量的增加,饲料适口性较差,降低了黄姑鱼幼鱼的摄食率,从而降低了肝脏中的IGF-I的表达量。前人在军曹鱼幼鱼Rachycentron canadum[26]、花鲈Lateolabrax japonocus[27]和黄河鲤鱼Cyprinus carpio[28-29]饲料研究中发现植物蛋白添加比例较高时,肝脏中IGF-I的表达量较低,这和本实验结果是类似的。有趣的是在本实验中发现黄姑鱼肌肉中IGF-I表达量呈先上升后下降的趋势,在FSM30组时显著最高。这与肝脏中的IGF-I表达模式不同,分析原因可能与适量的发酵豆粕替代鱼粉后增加了黄姑鱼肌肉中氨基酸的沉积率有关,具体原因尚待进一步研究。
3.3 不同水平发酵豆粕替代鱼粉对黄姑鱼幼鱼肝脏组织形态的影响
过量的植物蛋白替代鱼粉会影响肝功能,当肝脏发生病变,脂蛋白合成减少,使得肝细胞中的脂肪无法及时运出,造成肝脏脂肪堆积[30-31]。近年来,在对杂交鲟Acipenser baeri♂×A.schrenkii♀[32]、罗非鱼[33]、大黄鱼Larimichtys crocea[34]植物蛋白替代鱼粉的研究中发现随着混合植物蛋白替代鱼粉比例升高,肝脏逐渐出现病变,当植物蛋白替代鱼粉比例过高时,肝细胞空泡化严重。本实验中黄姑鱼肝脏组织学观察显示,FSM0组细胞排列规则整齐,细胞核正常;FSM20组的肝细胞出现少量空泡化;而FSM50组肝脏细胞空泡化现象严重,肝细胞轮廓模糊,部分肝细胞核已经消失不见。这与上述鱼种研究结果相似,过量的发酵豆粕的添加会影响肝功能,导致肝脏发生病变。因此,在实验中应注意发酵豆粕的添加量。
4 结论
本实验条件下,发酵豆粕替代鱼粉可以有效提高黄姑鱼鱼体中氨基酸的积累(除缬氨酸和蛋氨酸),替代量在20%~30%为宜,否则会降低肝脏和肌肉中IGF-I的表达量并造成肝脏损伤。
