亚低温对不同品种番茄幼苗干物质积累与氮磷吸收及分配的影响
2018-08-02吴凤芝周新刚高丹美杨万基
高 欢,吴凤芝, 周新刚,高丹美,杨万基
(东北农业大学 园艺园林学院,哈尔滨 150030)
番茄起源于热带,对低温反应敏感[1],是设施栽培的主要蔬菜。由于我国温室的结构相对简陋以及大多数的生产者为了节约成本,因此较少或不采用加温设备,番茄在北方冬春季设施生产中常常遭遇亚低温胁迫[2]。亚低温对蔬菜的生长发育、营养元素的吸收分配、干物质积累以及产量和品质均会产生巨大影响[3-4]。研究不同耐低温番茄品种在亚低温条件下养分吸收分配及干物质积累特征,对冬春季设施番茄的施肥管理以及提高设施番茄的产量和品质均具有重要的意义。
关于亚低温对植株生长发育的影响,前人已有大量研究。亚低温明显抑制了植株的生长,使番茄幼苗株高和叶面积显著减小,干物质积累下降[5],番茄种子的发芽率降低[6],叶片厚度变薄,栅栏组织厚度减小[7],果实品质下降[8]。亚适宜根区温度阻碍矿质元素的吸收与分配[9]。研究表明,植株体内磷和钾的浓度与积累量均受到根区温度的显著影响[10-12];黄瓜结果期处于亚适宜温光条件下,根系、叶片与果实中氮含量与氮转运蛋白基因的表达均下降[13];不同抗性品种在低温条件下的生理特性不同,低温胁迫后,抗寒性强的小麦光合机构受损伤的程度轻,具有较高的光合活性和较强的自我保护机制[14]。孙守如等[15]研究指出,耐寒性较强的西葫芦品种在低温条件下根系活力与干物质积累量均大于冷敏品种。总的来看,关于亚低温对番茄幼苗生长发育以及营养元素的吸收分配的影响,前人均已进行过研究,但针对耐寒性不同的2个番茄品种在亚低温条件下干物质积累以及养分吸收分配特征的研究甚少。本试验以耐寒性不同的2个番茄品种为试材,通过分析亚低温条件下番茄幼苗各器官中干物质与营养元素的积累与分配特性,为揭示不同番茄品种对亚低温条件的适应特征及其生长发育的调控提供理论依据。
1 材料与方法
1.1 供试材料
供试的番茄品种为耐寒品种‘东农722’与冷敏品种‘东农708’,由东北农业大学番茄研究所提供,供试基质为蛭石与珍珠岩,订购于哈尔滨美莱生物科技有限公司。
1.2 试验方法
1.2.1 试验设计 试验于2016年8-11月在东北农业大学设施生理生态研究室进行。种子消毒后于55 ℃温水中浸泡10 min,然后将种子放于清水中浸种6 h,放于28 ℃恒温培养箱中催芽,待70%种子露白后播种于育苗盘中,育苗用土为大田土。待番茄幼苗长至2叶1心时,分苗于装有基质的塑料营养钵(10 cm×10 cm)中,每钵1株,基质由蛭石和珍珠岩按照体积比2∶1配制而成。分苗后采用半剂量改良的Hogland营养液进行浇灌,每3 d浇1次营养液,每钵浇100 mL。待幼苗长至4叶1心时,选取整齐一致的番茄幼苗移至光照度12 000 lx,相对湿度65%的人工气候箱(HPG-280HX)中进行处理。温度设置为适温25 ℃/18 ℃(昼/夜)与亚低温15 ℃/10 ℃(昼/夜) 2个水平,光周期均为16 h/8 h (昼/夜)。设4个处理:分别以适温条件下的耐寒番茄品种‘东农722’(DN-722 25 ℃/18 ℃)和冷敏番茄品种‘东农708’(DN-708 25 ℃/18 ℃)为对照,设亚低温条件下的耐寒番茄品种‘东农722’(DN-722 15 ℃/10 ℃)、冷敏番茄品种‘东农708’(DN-708 15 ℃/10 ℃)。每个处理60钵,重复3次,随机排列。
1.2.2 测定项目及方法 分别于亚低温处理后的第5天和第10天对各处理选取整齐一致的番茄幼苗9株,洗净后吸干水,然后切分为根、茎和叶3部分,称鲜质量后放入烘箱内在105 ℃下杀青30 min,然后于65 ℃条件下烘干至恒量,用万分之一天平称量干质量。将烘干称量后的样品用粉碎机粉碎,粉碎后的植株过0.3 mm筛,用浓H2SO4消煮,消煮液用于养分的测定。根、茎、叶中氮和磷质量分数均采用SKALAR流动分析仪进行测定。
1.3 数据分析
试验中原始数据的整理采用Excel 软件完成,采用SAS 9.1软件进行数据分析,多重比较采用Tukey’s HSD法(P<0.05)。
2 结果与分析
2.1 亚低温对番茄幼苗干物质积累与分配的影响
2.1.1 对番茄幼苗干物质积累的影响 处理5 d后,亚低温(15 ℃/10 ℃)显著降低了DN-708与DN-722番茄幼苗总干质量、地上部干质量及茎与叶干质量(图1-A~1-E),总干质量分别降低了30.95%和22.22%;处理10 d后,与对照(25 ℃/18 ℃)相比,DN-708根、茎和叶干质量的下降幅度分别为14.29%、29.63%和21.05%,低于DN-722的28.57%、41.46%和24.07%(图1-C~1-E);2个品种相比,相同处理条件下,耐寒品种DN-722各器官中的干质量均显著高于冷敏品种DN-708(图1-C~1-E),处理5 d和10 d后,DN-722亚低温条件下的总干质量分别是‘东农708’的2.0和1.3倍(图1-A)。
2.1.2 亚低温对番茄幼苗各器官干物质分配的影响 番茄幼苗各器官中干物质的分配率表现为叶>茎>根,亚低温(15 ℃/10 ℃)增大了番茄幼苗根中干物质的分配率并降低了茎中干物质的分配率。与对照(25 ℃/18 ℃)相比,2个品种番茄根中干物质分配率在处理5 d后增加的幅度相同,均为2.2%;处理10 d后,DN-708叶中干物质分配率增加的幅度为1.3%,低于DN-722的5.2%;处理5 d和10 d后,DN-708茎中干物质分配率与对照相比下降幅度分别为2.5%和4.0%,低于DN-722的4.3%和5.9%(图2)。
2.2 亚低温对番茄幼苗各器官氮质量分数及其积累与分配的影响
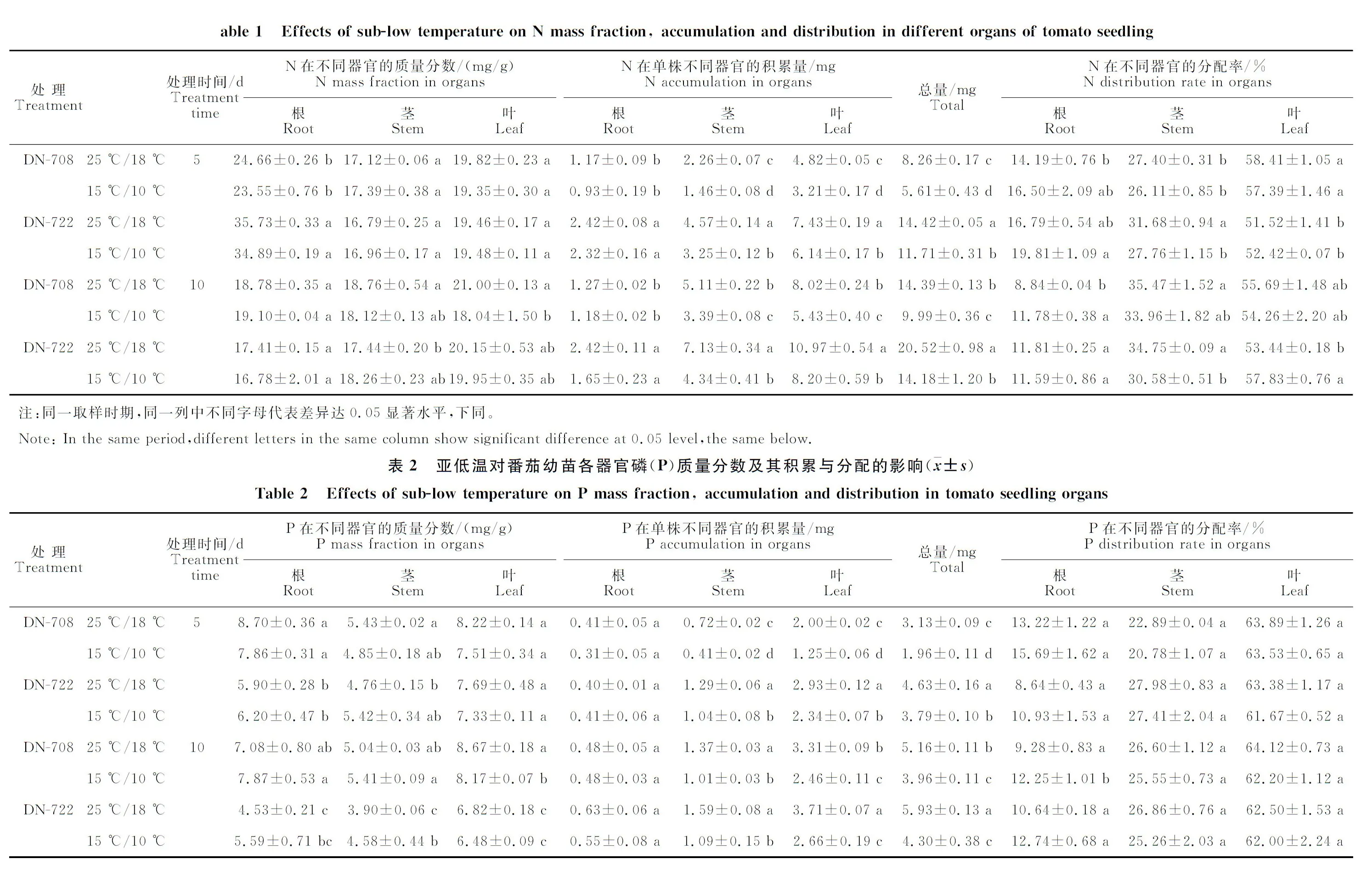
由表1可见,亚低温(15 ℃/10 ℃)处理10 d后,DN-708叶中的氮质量分数与对照(25 ℃/18 ℃)相比下降了14.10%,达到差异显著水平。氮在各器官中的积累量表现为叶>茎>根,与对照相比,亚低温显著降低了番茄幼苗单株氮积累量与茎、叶中氮的积累量。处理5 d和10 d后,DN-708与DN-722单株氮积累量与对照相比分别下降32.08%、30.58%与18.79%、30.89%。DN-708根与DN-722叶中氮的分配率在处理10 d 后与对照相比显著增加。2个品种相比,处理 5 d 后,亚低温条件下耐寒品种DN-722根中氮的质量分数显著高于冷敏品种DN-708,而叶中氮的分配率显著低于DN-708;处理5 d和10 d后,DN-722 各器官中氮的积累量在相同处理条件下均显著高于DN-708 ,亚低温条件下的单株氮积累量是DN-708的2.1倍,处理10 d后则为1.4倍。
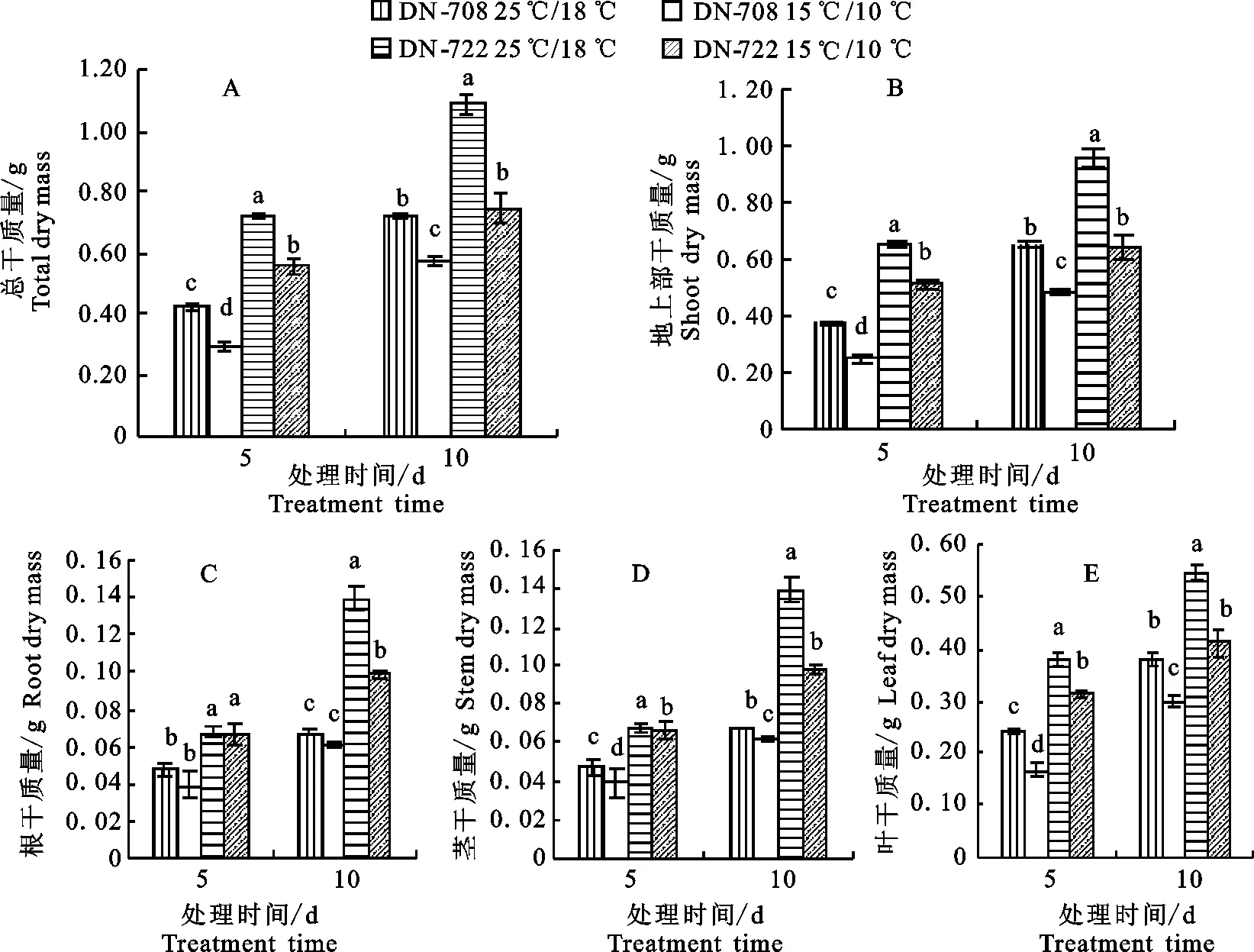
同一时期柱形图上不同字母代表差异达0.05显著水平 In the same period, different letters above the column show significant difference at 0.05 level
图1亚低温对番茄幼苗总干质量(A)、地上部干质量(B)、根干质量(C)、
茎干质量(D)和叶干质量(E)的影响
Fig.1Effectsofsub-lowtemperatureontotaldrymass(A),shootdrymass(B),rootdrymass(C),
stemdrymass(D)andleafdrymass(E)oftomatoseedling

图2 亚低温对番茄幼苗各器官干物质分配特性的影响Fig.2 Effects of sub-low temperature on dry matter distribution rate in different organs of tomato seedling
2.3 亚低温对番茄幼苗各器官磷质量分数及其积累与分配的影响
表2显示,亚低温(15 ℃/10 ℃)处理10 d后,与对照(25 ℃/18 ℃)相比,DN-708叶中磷质量分数降低了5.8%,DN-722茎中的磷质量分数增加了17.44%,均达到差异显著水平。DN-708与DN-722单株磷积累量在处理5 d后与对照相比分别下降了37.38%与18.14%,处理10 d后则分别下降了23.26%与27.49%;2个品种相比,处理5 d 后,相同处理条件下,均表现为耐寒品种DN-722单株磷积累量与茎和叶中的磷积累量显著高于冷敏品种DN-708,且亚低温条件下单株磷积累量是‘东农708’的1.9倍;处理10 d后,‘东农708’各器官中的磷质量分数均显著高于DN-722,根中磷的分配率在亚低温条件下低于DN-722,茎与叶中磷的分配率2个品种间无显著差异(表2)。
3 讨 论
在植物生长发育过程中,温度作为一个重要的环境因子对植物的生长发育起着至关重要的作用[16]。对于喜温的番茄来说,随着温度降低,生长迅速减慢[17]。本研究结果表明,亚低温(15 ℃/10 ℃)显著降低了番茄幼苗茎与叶中干物质积累量,根中干物质积累量在处理5 d后没有明显的下降。这与Alfandi[18]在甜椒上的研究结果相似,其得出的结果也表明,亚低温对根生长的影响不大,而茎和叶的生长则比较缓慢。这可能是由于低温对叶片线粒体的伤害大于根系,番茄幼苗地上部与根系对低温的敏感性可能与呼吸作用相关[19]。
本试验结果发现,处理10 d后,耐寒品种DN-722 茎与根中干物质的积累量下降的幅度高于冷敏品种DN-708,同时叶片中的干物质与氮的分配率增加的幅度大于DN-708,可能是为了维持植株正常生长与光合作用,DN-722将大多数的营养分配到了叶片中,从而导致其茎和根中干物质的积累量下降幅度相对较大。番茄光合产物主要通过光合作用的强弱来体现,低温抑制番茄的光合作用,进一步抑制总干物质的合成[20]。王国莉等[21]研究表明低温条件下冷敏品种光合速率的下降幅度大于耐寒品种。不同耐寒性番茄品种在亚低温条件下光合特性的不同,可能是导致干物质的积累量存在差异的主要原因。温度影响作物的库强,干扰作物干物质分配。从本试验可以看出,亚低温增大了DN-708与DN-722根中干物质的分配率,降低了茎中干物质的分配率,这可能是由于亚低温条件下,为了保持植株的根系活力和维持植株的存活,需要增加根系功能单位来修复或降低逆境的影响,光合产物较多的分配到根系,从而抑制地上部的生长。也有研究表明,亚低温条件下,干物质向茎与叶中的分配比率均增加,与本试验的结果不同,可能是亚低温的处理时间和温度设置不同导致的[22]。
在蔬菜生命周期的各阶段,土壤氮、磷和钾等矿质营养元素的稳定供应是保证蔬菜正常生长发育的前提[23]。温度降低大多会降低矿质离子的主动吸收[24]。Tachicana等[25]在黄瓜上的研究结果表明,温度低于17~20 ℃时,黄瓜对水分与矿质营养的吸收和运转均受到抑制。本试验中,番茄幼苗根中磷质量分数在亚低温条件下有增高的趋势,而叶片中的氮和磷质量分数则有降低的趋势,这与白青华[23]的研究结果一致。两品种比较,亚低温条件下,冷敏品种 DN-708各器官中磷质量分数均大于耐寒品种DN-722,可能是由于DN-722干物质的积累量增加的幅度大于磷增加的幅度,因而单位干质量内的磷质量分数较低[26],因此,不能仅仅通过植株体内矿质元素的吸收和积累量来判断番茄的耐寒性强弱,还应该考虑与产量相关的其他生长发育指标。
矿质元素的积累与干物质的积累密切相关,随着干物质的积累,矿质元素在植物体内不断增加,植物对矿质元素的积累又是干物质积累的基础[27-28]。根区温度是重要的环境因子,对植物生长发育所需的矿质营养元素的有效浓度产生重要的影响[29-30]。本试验中,亚低温处理条件下,番茄幼苗茎与叶中氮、磷积累量显著下降,这与刘玉凤等[31]的研究结果一致。主要原因可能是亚低温条件下根系组织结构破坏,降低了对营养元素的吸收,从而使植株体内营养元素含量降低,也可能是由于亚低温降低了番茄根系营养元素吸收转运相关基因的表达量,从而抑制了营养元素向地上部的转运。2个品种相比,对照与亚低温条件下,耐寒品种DN-722各器官中氮的积累量均大于冷敏品种DN-708,而茎和叶中的磷积累量在亚低温处理5 d 时大于冷敏品种DN-708。这可能是亚低温条件下,不同耐寒性番茄对营养元素的利用效率以及吸收转运调控机制不同导致的。温度影响干物质与矿质营养向番茄各部位的分配比例[32]。矿质元素在植物体各器官中的分配比例反应该元素在植株体内的分布及其在各器官间的迁移规律[33]。从本试验可以看出,亚低温对2个品种番茄幼苗各器官中磷分配率影响不大,而显著增加了DN-722叶与DN-708根中的氮分配率,耐寒品种DN-722为了维持正常的生长,从而使氮向叶片中的分配增多,而冷敏品种DN-708则可能是转运能力受到抑制,因而根中滞留的氮较多。
4 结 论
亚低温条件下,番茄幼苗干物质及氮和磷积累量均显著下降。两品种比较,耐寒品种单株干物质积累量与氮和磷积累量均高于冷敏品种, 且在亚低温处理5 d后以上3个指标下降的幅度均小于冷敏品种,处理10 d后下降的幅度则均高于冷敏品种。随着处理时间的延长,番茄幼苗根与叶中干物质与氮的分配率均增加,而茎中两者的分配率则降低。总的来看,耐寒品种受亚低温的影响迟于冷敏品种,可为冬春季的栽培管理提供一定的参考依据。
