黄瓜氮素胁迫相关基因CsAHP1的克隆及功能分析
2018-08-02杜亚琳陈海燕徐丽琳范莲雪
杜亚琳,陈海燕,徐丽琳,王 琛,范莲雪
(1.农业部东北地区园艺作物生物学与种质创制重点实验室/东北农业大学 园艺园林学院,黑龙江 哈尔滨 150030;2.河南省农业科学院 农业经济与信息研究所,河南 郑州 450002)
氮是植物生长发育的重要营养元素之一。但过量施用化肥,特别是氮肥,是目前黄瓜等农作物生产过程中面临的主要问题。据报道,化肥中所包含的氮只有30%~50%被植物吸收和利用[1-3]。黄瓜是一种富含硝酸盐的蔬菜。近年来,黄瓜栽培过程中氮肥的过量施用已经导致环境严重污染[4],并且也抑制了黄瓜植株的生长发育[5]。此外,蔬菜硝酸盐污染是我国绿色蔬菜以及无公害蔬菜生产中面临的主要问题,而过量施用氮肥不仅是造成该问题的主要原因之一[6-9],还影响了农民的收入。因此,培育耐氮素胁迫能力强的黄瓜新品种对减轻环境受损及降低黄瓜生产成本意义重大。
细胞分裂素是参与调节植物生长发育过程的重要激素之一[10-13],而含有组氨酸的磷酸转移蛋白(AHPs)是细胞分裂素信号级联反应的重要组成部分,拟南芥中共有6种AHP蛋白[14]。然而在低氮胁迫条件下AHPs在黄瓜中的作用尚不清楚。
为深入了解黄瓜生殖生长过程中的氮循环与其耐氮素胁迫能力之间的关系,前期对黄瓜生殖生长期叶片的转录组进行分析,获得了1个黄瓜氮素胁迫相关候选基因CsAHP1(Csa1M572420)[15]。本研究对CsAHP1在不同氮素处理条件下的表达及其序列进行了分析,构建了35S∶∶CsAHP1拟南芥过表达转基因株系,并对转基因拟南芥植株耐氮素胁迫能力进行了分析,为深入理解CsAHP1基因参与黄瓜耐氮素胁迫的生理与分子机制提供参考。
1 材料和方法
1.1 材料种植及氮素处理
本研究以黄瓜津研四号为试验材料,对其进行低浓度氮素处理试验。将黄瓜津研四号种子先进行催芽,然后移植于装有蛭石的营养钵中,分别用3 mmol/L和14 mmol/L 的KNO3溶液进行处理,处理后37 d对幼苗进行全株取样。
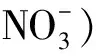
1.2 qRT-PCR分析


表1 基因克隆、表达分析和载体构建所需引物序列
注:下划线表示酶切位点。
1.3 CsAHP1基因的克隆及序列分析
在葫芦科基因组数据库(http://www.icugi.org/cgi-bin/ICuGI/index.cgi)中搜索CsAHP1基因ID号 Csa1M572420.1,获得其CDS序列信息,以该CDS序列为模板,使用Primer Premier 5.0设计克隆引物(表1),并由上海生物工程技术服务有限公司合成。利用PCR方法进行CsAHP1基因的克隆。利用NCBI(http://www.ncbi.nlm.nih.gov/)在线Blast工具和DNAMAN 8.0软件进行序列比对和分析,利用MEGA 6.0软件的邻近法进行系统进化树的构建。通过在线软件Expasy对该基因所编码的蛋白质理化性质进行预测分析。
1.4 CsAHP1过表达转基因拟南芥株系的获得
将CsAHP1基因的全长编码序列克隆到带有XbaⅠ/BamHⅠ酶切位点的pBI121载体中,然后将所构建好的重组体转化到农杆菌菌株LBA4404中,最后通过花序侵染法[18]转化到Col-0植株中。在含有卡那霉素(Kan)的MS培养基上筛选转化的种子(T1),并最终获得纯合的T3代种子,即35S∶∶CsAHP1转基因拟南芥株系。
1.5 拟南芥植株根长和质量的测定

2 结果与分析
2.1 黄瓜耐氮素胁迫基因CsAHP1的表达分析
本研究前期通过Solexa测序技术对黄瓜耐氮素胁迫相关基因的表达谱进行了分析,鉴定出1个在低氮素胁迫条件下上调表达的基因CsAHP1[15]。为进一步验证CsAHP1与耐氮素胁迫之间的关系,对CsAHP1的表达进行了qRT-PCR分析。结果表明,在低氮素胁迫条件下,CsAHP1基因表达水平显著增加,这一结果与前期Solexa测序结果[15]一致(图1),表明CsAHP1为一个黄瓜低氮素胁迫诱导表达的基因。
2.2 黄瓜CsAHP1基因的克隆及序列分析
黄瓜CsAHP1基因cDNA长462 bp,编码153个氨基酸。在线软件Expasy-ProtParam预测结果表明,CsAHP1蛋白分子质量为17.78 ku,理论等电点为5.76。NCBI数据库检索结果显示,CsAHP1属于HPT家族。序列分析表明,CsAHP1的氨基酸序列与甜瓜的同源性为95%,与巨桉和川桑的同源性为64%,与拟南芥、烟草和番茄也均有> 60%的同源性,表明CsAHP1基因在不同物种中高度保守(图2)。此外,在CsAHP1的同源序列中发现了HPT家族的典型特征,即组氨酸残基(图2)。
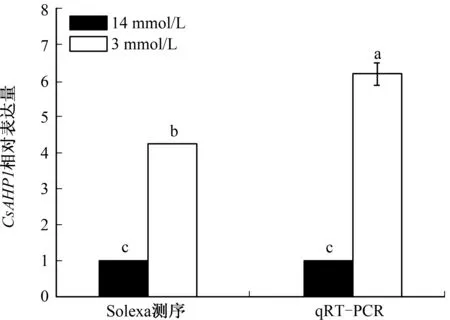
不同字母表示在0.05水平差异显著,下同图1 CsAHP1在不同氮素处理条件下的表达水平
为了解黄瓜CsAHP1和其他物种AHP蛋白之间的进化关系,利用MEGA 6.0进行进化树的构建,结果表明,黄瓜CsAHP1和甜瓜关系最近(图3)。
2.3 过表达CsAHP1基因拟南芥耐氮素胁迫能力分析
为进一步阐明CsAHP1参与耐低浓度氮素胁迫过程的生物学功能,本研究构建了CsAHP1基因过量表达载体,并转入了拟南芥植株,获得了1个35S∶∶CsAHP1转基因拟南芥株系。根长测定结果表明,在低浓度氮素和高浓度氮素生长条件下,转基因植株根长均显著大于Col-0植株(图4A、B)。在低浓度氮素条件下,Col-0植株的根长为1.34 cm,不到转基因植株幼苗根长的44.7%(图4B)。在高浓度氮素和低浓度氮素胁迫情况下,通过比较分析Col-0相对于转基因株系在根长方面的变化程度发现,Col-0植株的根伸长发育受抑制程度(56.2%)显著大于转基因植株(36.7%)(图4C)。

图3 CsAHP1蛋白及其同源蛋白的系统进化树
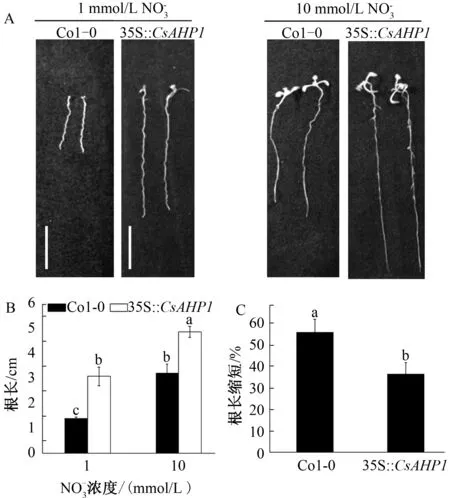
A:在高氮和低氮条件下生长10 d的拟南芥Col-0植株和35S∶∶CsAHP1过表达转基因植株;B:Col-0植株和转基因植株根长统计;C:低氮下Col-0植株和转基因植株根长的减少比例图4 不同氮素条件下的拟南芥植株
在低浓度氮素和高浓度氮素情况下,Col-0植株的质量分别为1.34 mg和16.53 mg;而转基因植株幼苗的质量分别为4.37 mg和19.83 mg(图5A)。相对于高浓度氮素条件下植株的质量,在低氮素浓度胁迫条件下Col-0植株质量下降程度(91.89%)显著大于转基因植株(77.96%)(图5B)。综上所述,CsAHP1的过表达显著提高了转基因植株的耐氮素胁迫能力。
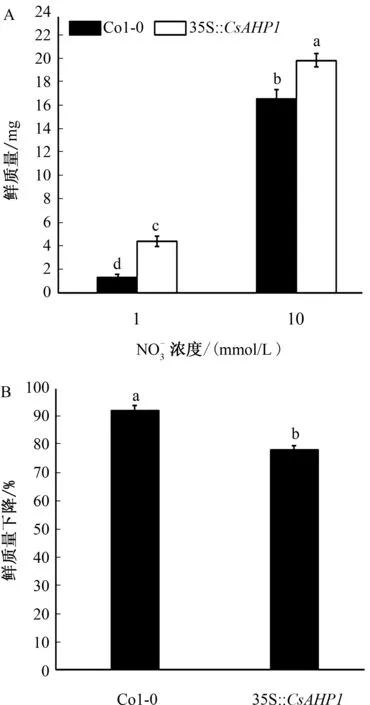

图5不同氮素条件下拟南芥Col-0植株和35S∶∶CsAHP1过表达转基因植株质量比较
3 结论与讨论
本研究在高浓度氮素及低浓度氮素胁迫条件下对拟南芥Col-0植株和35S∶∶CsAHP1过表达转基因拟南芥株系根长及质量的测定分析结果表明,35S∶∶CsAHP1过表达转基因植株具有比Col-0植株更强的耐低浓度氮素胁迫能力。在本研究中,对这些结果给出了几种可能的解释。其中一个假设是CsAHP1可能通过细胞分裂素信号通路参与耐低浓度氮素胁迫过程。细胞分裂素调节植物的生长发育[19-24],而AHP蛋白在细胞分裂素信号通路中起重要作用[25]。Yokoyama等[26]证明了CsAHP1的下游基因ARR10和ARR12参与拟南芥根的分化和伸长过程,而该过程受细胞分裂素调节。CsAHP1基因过表达可以使ARR10和ARR12出现上调表达,从而可能促进了细胞分裂素的合成以维持拟南芥植株的正常生长。有研究发现,外源细胞分裂素处理可缓解车前草因低浓度氮素胁迫所引起的生长抑制问题[27]。大多数在植株茎间中表达的AtNRTs都受细胞分裂素处理诱导表达,由此促进硝酸盐的运输和分布[28]。此外,低浓度氮素胁迫条件下拟南芥幼苗根部生长素含量极高[26]。细胞分裂素通过调节生长素的外排量来调节生长素的分布[29],生长素-细胞分裂素相互作用控制根分生组织的发育[30-31]。因此,CsAHP1还可能通过促进细胞分裂素合成,调节生长素分布以提高根分生组织活力,进而增强其耐低浓度氮素胁迫能力。
