青藏高原高寒栽培草地植物在光照变化和增温影响下矿物元素蓄积分异性初探*
2018-07-30李天才曹广民张吉余郭小伟
李天才, 曹广民, 林 丽, 张吉余, 郭小伟, 柯 浔
(中国科学院西北高原生物研究所,青海省藏药药理学和安全性评价研究重点实验室,青海 西宁 810008)
矿物元素是植物生长发育的必需营养,而光照和温度也是植物生长发育必要的环境影响因素。退化草地较封育草地植物中常量矿物元素和微量元素含量均有增加,即退化草地植物中矿物元素具有蓄积分异性,生物矿物元素饥饿效应是草地矿物元素蓄积分异行为发生的内动力,草地植物中矿物元素营养的变化也是草地植被演替的重要响应[1-3]。在青藏高原高寒、干旱和强紫外辐射等恶劣环境下,光照和温度等生态因子也是草地矿物元素蓄积分异的重要影响因子[4-17]。本研究通过天然高寒草地中优势种植物垂穗披碱草和青海冷地早熟禾盆栽试验,模拟增温和光照强度变化等环境影响因子影响下其中矿物元素营养的变化,说明在光照和温度影响下草地植物中矿物元素的变化趋势,解释高寒草地植物中矿物元素的蓄积分异机制,研究对于退化草地中矿物元素蓄积分异机制研究,生物矿物元素饥饿效应假说理论的检验等具有重要意义,也将为退化草地恢复与修复,天然草地保护、草地生产力提高等提供一定的科学依据和技术支撑。
1 材料与方法
1.1 试验材料
垂穗披碱草(Elymusnutans):多年生禾草。根茎疏丛状,须根发达,茎叶茂盛。根茎分蘖能力强,抗寒,能安全越冬。适应性强,对土壤要求不严,各种类型的土壤均能生长。抗旱力较强,不耐长期水淹。喜生长在高原滩地以及山地阳坡、沟谷、半阴坡等地。草质柔软,无刺毛、刚毛,无味,从返青到开花前,马、牛、羊最喜食。调制的青干草是冬春季马、牛、羊的良好保膘牧草。属中上等品质牧草[14]。
青海冷地早熟禾(Poacrymophila):疏丛型禾草,茎杆直立,茎叶茂盛。根茎发达,分蘖能力强。能适应高原复杂的生境条件,无论海拔高低均能良好生长。抗旱、耐盐碱、耐瘠薄、抗寒,对土壤要求不严格。人工栽培的冷地早熟禾草地可作为割草场,或割草与放牧兼用[14]。
青海冷地早熟禾茎杆柔软,略带甜味,适口性好,枝叶茂盛,营养枝发达营养生长期较长。籽实成熟后枝叶仍保持青绿,叶片不易脱落,各个不同发育期粗蛋白含量接近苜蓿,开花前马、牛、羊最喜食,干草质地柔软,可作为冬春的良好补充饲料,是夏秋各类家畜的抓膘优良牧草,冬春则是救命保膘草。
垂穗披碱草和青海冷地早熟禾均为适宜青藏高原栽培种植的草地植物,有市售商品化种子(试验用垂穗披碱草和青海冷地早熟禾种子均由青海省畜牧兽医科学研究院草原研究所德科加研究员提供),易于种植,生长期较长,为草地植物试验研究比较理想的草地植物物种。
1.2 盆栽试验
试验于2016年4月中旬于西宁(中国科学院西北高原生物研究所院内)盆栽垂穗披碱草和青海冷地早熟禾各30盆(盆径约30 cm),盆栽土壤用草地黑壤土(采自达坂山海北高寒草甸定位站周边牧场,混有牛羊粪自制的有机肥),垂穗披碱草和青海冷地早熟禾的播种量分别为2.0 g·m-2、1.0 g·m-2。试验主要设计为两个处理,分别为遮阴和模拟增温,并各自设有对照。
A为遮阴组:以工程用遮阴网布为遮阴材料,自制试验用遮阴架,生长期视植物生长高度,适时调整遮阴网高度,试验组白天平均光照强度约为对照组平均光照强度的53.4%,灌水量同对照组,灌水周期视天气状况为1次/ 1~2天。
B为增温组:以农用塑料覆膜为增温材料,自制试验用增温架,生长期视植物生长高度,适时调整增温网高度,试验组白天温度相对于对照组平均增加约2℃,灌水量同对照组,灌水周期视天气状况为1次/ 1~2天。
试验持续至8月下旬的牧草成熟期,茎叶尖略枯黄,高约为30~40 cm,尚未抽穗结子(多年生植物,盆栽第一年),即生长期为4个月,按试验要求各自采样,阴干,备用。将采集的各试验草地植物样品,经自然阴干、去离子水清洗、60℃下烘干、研磨、过100目筛等样品预处理后,送中国科学院西北高原生物研究所分析测试中心进行矿物元素分析测试。
1.3 分析测试
分析仪器:电感耦合等离子质谱仪(NexIONTM 350D,美国PerkinElmer公司)。
分析元素:Ca、Mg、K、Na、P 、Cu、Zn、Fe、Mn、Cr共10种元素。
1.4 数据分析
采用SPSS 20.0 统计学软件对各组数据进行单因素方差分析。
2 试验结果
2.1 光照试验结果
不同光照条件(遮阴试验)下,盆栽垂穗披碱草和青海冷地早熟禾中矿物元素含量,见表1与表2。
在本次选取的样本当中,二维超声共出现6例漏诊案例,三维超声出现1例漏诊案例,三维超声诊断的符合率也要高于二维超声。

表1 不同光照下盆栽垂穗披碱草与青海冷地早熟禾中矿物常量元素Table 1 The mineral macroelements of cultivated Elymus nutans and Poa crymophila under different light
注:与对照组比较,*为P<0.05,**为P<0.01,下同
Note: Comparing with control group, *indicates difference at the 0.05 level;**indicates difference at the 0.01 level, the same as below

表2 不同光照下盆栽垂穗披碱草与青海冷地早熟禾中微量常量元素Table 2 The mineral trace elements of cultivated Elymus nutans and Poa crymophila under different light
2.2 增温试验结果
不同温度条件(增温试验)下,盆栽垂穗披碱草和青海冷地早熟禾中矿物元素含量,见表3与表4。
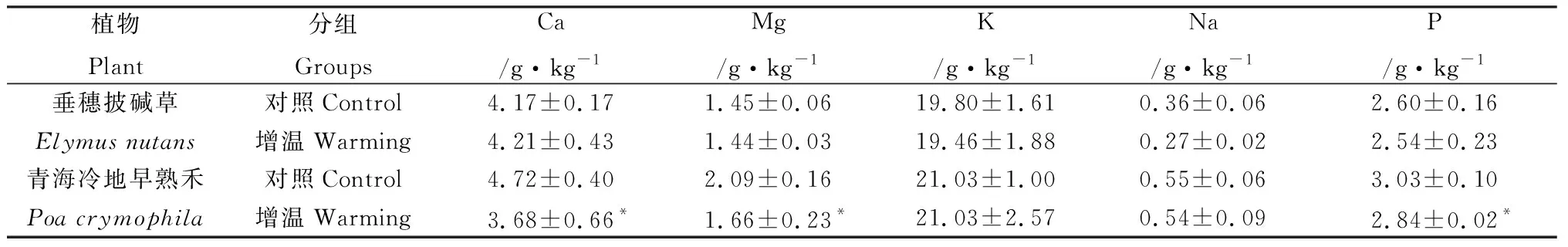
表3 不同温度下盆栽垂穗披碱草与青海冷地早熟禾中矿物常量元素Table 3 The mineral macroelements of cultivated Elymus nutans and Poa crymophila under different temperatures
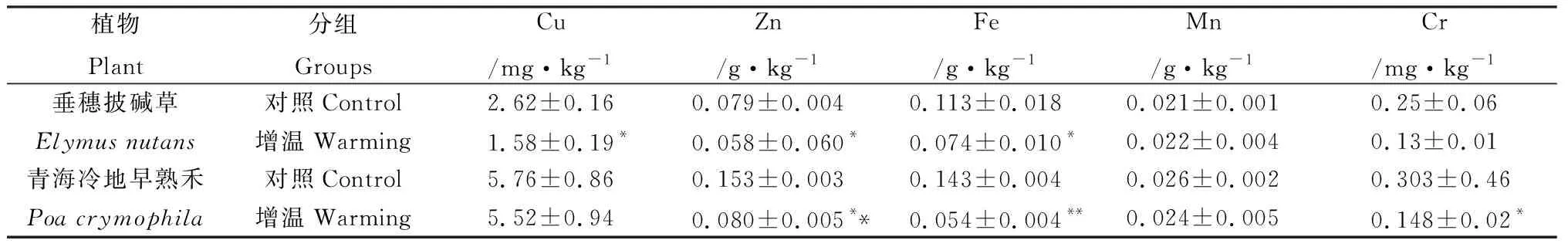
表4 不同温度下盆栽垂穗披碱草与青海冷地早熟禾中微量常量元素Table 4 The mineral trace element of cultivated Elymus nutans and Poa crymophila under different temperatures
3 讨论
3.1 不同光照条件下盆栽草地植物中矿物元素蓄积性
由表1与表2,光照强度对盆栽草地植物中矿物元素有较明显的影响,其中垂穗披碱草中除P元素外,遮阴组较对照组矿物元素含量均有增加,而青海冷地早熟禾中除元素Cu、Zn外,遮阴组较对照组矿物元素含量也是在增加,且盆栽两草地植物中矿物元素Na、Zn和Mn表现出极显著的差异性(P<0.01),元素Ca、Mg具有显著的差异性(P<0.05)。
植物对土壤或介质中矿物元素养分的吸收具有:选择性、累积性和基因型差异[4]。由表1与表2可知,盆栽垂穗披碱草和青海冷地早熟禾对于土壤环境中同一矿物元素的吸收表现为对不同矿物元素营养的选择性和植物种的差异性,即基因型差异。因此,在相同的光照和温度条件下,盆栽垂穗披碱草和青海冷地早熟禾对于同一矿物元素营养的吸收量和累积量不同。如:同样环境条件下,对照组垂穗披碱草和青海冷地早熟禾中Cu分别为2.52 mg·kg-1和5.76 mg·kg-1,青海冷地早熟禾中Cu的累积量是垂穗披碱草中Cu的累积量的1.29倍。
光照强度对植物的生长、发育及形态结构的建成都有重要作用[5]。当其他环境条件不变的情况下,光照强度的变化决定着净光合作用的变化,光照不足将引起植物体内养分供应的障碍,可导致已形成的花芽、果实发育不良或早期死亡,也会影响果实的品质[5]。试验中遮阴组较对照组草地植物中矿物元素的蓄积增加,表明光照强度对草地植物的生长发育极为重要,且表现出植物中矿物元素含量随光照强度的减小而蓄积增加的特征。
矿物元素也是植物生长必须的营养物质,元素Mn参与光合作用,与叶绿体合成有关,是维持叶绿体结构所必需的元素,在光合作用中Mn参与水的光解[6]。当在遮阴减小光照强度时,植物的净光合作用发生变化而减弱,引起植物体内养分供给的障碍。遮阴组相对于对照组植物,矿物元素Mn供给因光照而受影响,即遮阴组植物体内矿物元素Mn营养因供给障碍不能满足植物生长的生理需求,处于对矿物元素Mn营养的“饥饿”状态,也就是说,遮阴组植物体内矿物元素Mn的蓄积增加就是生物矿物元素“饥饿效应”现象的表现。也即是遮阴组植物对于矿物元素Mn营养的“饥饿”而使体内的Mn元素蓄积增加,以适应因光照强度变化的新生长环境,也可以说,植物体内矿物元素Mn营养的蓄积增加是植物对光照强度减弱的生长环境变化的响应与适应。可见,生物矿物元素饥饿效应可能是遮阴组植物体内矿物元素蓄积分异行为发生的内动力之一,遮阴处理的光照强度变化试验也是对生物矿物元素“饥饿效应”假说理论的再检验。
矿物元素镁Mg也是叶绿素的成分,也是光合作用及呼吸作用中许多酶的活化剂,镁也是染色体的组成成分,在细胞分裂过程中起作用[6]。与Mn元素营养一样,植物因光照强度发生变化而使Mg元素营养供给受影响时,遮阴组植物同样因Mg元素营养的“饥饿”而蓄积增加,以适应因光照强度变化的新生长环境。即植物体内矿物元素Mg营养的蓄积增加也是植物对光照强度减弱生长环境变化的响应与适应。
矿物元素是植物生长发育的必需营养物质。光照强度变化的遮阴试验中,其它矿物元素的蓄积增加,同样是因为光照强度的变化引起植物对矿物元素营养需求的“饥饿效应”所致,因此,盆栽草地植物在遮阴试验中多种矿物元素的蓄积增加正是生物矿物元素“饥饿效应”现象的表现,也是生物矿物元素“饥饿效应”假说理论的试验检验,同时,试验也说明高寒草地植物体内矿物元素营养的蓄积增加是草地植物对生长环境变化的响应与适应,而生物矿物元素饥饿效应可能是矿物元素蓄积分异行为发生的内动力之一。
3.2 不同温度条件下盆栽草地植物中矿物元素蓄积性
植物的正常生长要在一定的温度范围内(一般0~35℃)才能进行,在此温度范围内,随着温度的升高,植物生长加快[5]。可见,增温组较对照组植物的生长速度快,盆栽试验中增温组较对照组的出苗时间短(提前1~2天)、同期增温组植物生长的株高较对照组平均高约5 cm。由于植物的快速生长相应需要相对更多的必需矿物元素营养,即增温组较对照组盆栽植物对矿物元素营养的需求量相对更多。试验中增温组植物因为能够及时得到所需矿物元素营养,按照生物矿物元素饥饿效应假说,增温组相对于对照组植物体内的矿物元素含量会低一些,也即是增温组因环境温度的增加而更有利于摄取必需的矿物元素营养,而对照组相对于增温组来说因环境温度的制约,对矿物元素营养的需求受限,处于对矿物元素营养的“饥饿”状态,可见,盆栽增温试验中对照组矿物元素的蓄积分异性也是生物矿物元素“饥饿效应”现象的又一表现。
土壤温度不但影响根系的生理生化活性,也影响根系中水的移动性。在一定的温度范围内,随土壤温度提高,植物根系中水运输加快,反之则减弱[5]。增温组相对于对照组植物根系的生理生化活性较强,根系中水运输相对也快,相应增温组植物对矿物元素的需求量增加,适当的环境温度增加能够满足植物对矿物元素营养的需求,即对照组相对于增温组植物因环境温度稍低,对于矿物元素营养的需求不能及时满足其需求,处于相对“饥饿”状态而蓄积增加,即是增温试验中对照组矿物元素的蓄积分异也是生物矿物元素“饥饿效应”现象的一种表现。
从植物蒸腾作用来看,蒸腾作用可以促进植物生长发育,蒸腾作用引起木质部的上升液流,有助于根部吸收的无机矿物离子以及根中合成的有机物转运到植物体的各部分,满足生命活动需要[5]。又因为植物的气孔开度一般随温度的升高而增大。增温组相对于对照组植物的蒸腾作用增强,相应对矿物元素营养需求增加,而体内矿物元素却少于对照组,正是增温组植物因温度增加,相对于对照组更能够及时得到矿物元素营养需求而处于“未饥饿”状态,即从植物的蒸腾作用看,增温组植物中矿物元素含量的减少也是其对环境温度增加的适应与响应,是生物矿物元素“饥饿效应”假说理论的再检验。青藏高原草地生态系统中,草地植物中矿物元素随着海拔高度的增加而增加,可能与随着海拔高度增加而环境温度的降低有关。
在增温和遮阴试验中,盆栽草地植物对环境温度和光照强度的变化都很敏感,都能够引起植物体内矿物元素营养的蓄积增加。在干旱和水胁迫试验,喷施矿物元素营养液体肥试验中,同样表现出盆栽草地植物中矿物元素营养的蓄积增加[16-17]。试验中环境温度、光照强度、水分和矿物元素营养供给等环境影响因子的变化都能使草地植物中矿物元素营养蓄积增加,其共同的特征都是盆栽植物对于矿物元素营养供给变化的及时反映,结合草地生态系统中,退化草地植物较封育草地植物中矿物元素蓄积性,因此,草地植物中矿物元素营养的蓄积是其对生长环境变化的适应与及时响应,其实质都是对矿物元素营养供给变化的响应,即生物矿物元素“饥饿效应”是驱使草地植物中矿物元素蓄积分异行为发生的内动力之一。由于植物对环境介质中矿物元素营养的吸收具有选择性,同时植物对于环境介质中矿物元素的吸收具有基因型差异,因此,不同草地植物对各种矿物元素营养蓄积的机制各异,草地植物对于某一矿物元素营养蓄积分异性的生物学机制有待进一步研究。
4 结论
光照强度和温度对高寒草地植物中矿物元素营养的蓄积分异性均有一定的影响,其中遮阴减小光照强度会使草地植物中矿物元素蓄积增加,温度降低的对照组相对于增温组草地植物中矿物元素也蓄积增加。
在不同光照强度和温度变化条件影响下,盆栽高寒草地植物中矿物元素蓄积分异性是生物矿物元素饥饿效应假说理论的再检验,其中生物矿物元素饥饿效应是草地植物中矿物元素蓄积分异行为发生的内动力之一。
遮阴(光照强度)和增温(温度)试验也说明高寒草地植物体内矿物元素营养的蓄积增加是草地植物对生长环境变化的适应与响应。
