生物遮阴对银杏幼苗生长的影响
2018-07-27钱龙梁曹福亮汪贵斌
钱龙梁 ,薛 源 ,曹福亮 ,2,李 柠 ,李 强 ,汪贵斌 ,2
(1.南京林业大学 南方现代林业协同创新中心,江苏 南京 210037;2. 江苏省农业种质资源保护与利用平台,江苏 南京 210014)
光合作用对植物生长及其生理变化至关重要[1],而光合作用又离不开光合时间、光合面积、光合能力以及光合产物的分布和消耗这几大主要影响因素[2]。在夏季晴天、自然光照条件下,植物有明显的“光合午休”现象,净光合速率有所降低[3]。光照与植物的生物量有直接关系,有研究表明在80%遮阴处理下核桃的淀粉和可溶性糖含量均显著提高[4]。实际上,通过适当地调节光合作用效率有助于促进植物生长及其产量的提高。银杏叶片中含有很高的黄酮类和内酯类化合物,1965年,德国Willmar Schwabe医生将银杏叶提取物(EGb761)引入医学临床[5]。因此近些年来,国内外对银杏叶用林高度重视,并兴建了大面积的叶用林。我国早在19世纪初期就已建立了银杏叶用林,也对叶用林的栽培技术方面进行了大量的研究及探索[6]。然而,就银杏的光合作用对其生理生态方面的影响鲜有报道,另外,遮阴处理对银杏生长及其光合作用影响的研究甚少,而对银杏叶用林采用生物遮阴研究更是少之又少。本文通过研究生物遮阴对银杏幼苗生长及其光合作用的影响,从而为大面积推广银杏叶用林兴建和管理以及合理利用土壤资源提供一定的理论基础。
1 试验地概况
试验地位于安徽省宿州市(东经118°10′,北纬34°38′),该地区属于北温带季风气候,宿州冬季比较长,有4个月之久,比较寒冷;夏日炎热,持续时间较春秋两季短,气候适宜,雨水适中。年平均气温为15.7 ℃,年降水量为817.28 mm左右,梅雨持续时间较长。土壤为沙壤土,pH为7.2~ 8.0。
2 材料与方法
2.1 试验材料
2017年3月份在播种过银杏种子的大田中选择生长一致且健壮的1年生银杏实生苗的实验样地12块,样地规格为50 m×0.6 m。
2.2 试验设计
2017年6月8 日在选好银杏幼苗试验地的同一侧按照不同距离点播一排玉米种子,全光照的试验地不种植玉米,待玉米长成植株后形成不同程度的遮阴处理,本次采用4个实验处理,即全光照(CK,遮阴程度0%),玉米株间距10 cm(L1,形成遮阴程度约70%),株间距20 cm(L2,形成遮阴程度约50%),株间距30 cm(形成遮阴程度约30%),遮阴期间进行常规管理。每个区组间空一块地作为隔离带,每个处理3个重复,每块试验地面积30 m2,栽400株,试验采用完全随机区组试验。
2.3 统计分析
试验数据采用Excel 2003和SPSS 19.0软件进行方差分析、多重比较和相关性分析[7]。
2.4 指标测定及方法
2.4.1 生长指标测定
在8月末试验结束时每个处理随机选取10株幼苗,用游标卡尺(0.02 mm)测量地径,用钢尺(0.5 mm)测量苗高和主根长,整株挖出清洗干净后,按茎、叶、根3部分分开,分别测量鲜质量,然后在105 ℃杀青半小时后在80 ℃下烘干至恒质量,用分析天平分别测定其干质量。计算比叶面积(总叶面积/叶质量)、根生物量比(根生物量/总生物量)、茎生物量比(茎生物量/总生物量)、叶生物量比(叶生物量/总生物量)、根冠比(地上部分生物量/根生物量)[8]。
2.4.2 叶绿素含量测定
选取测定光合数据的功能叶片,剪碎,称鲜质量,在黑暗中用10 mL 95%的乙醇溶液于试管中浸提至叶碎片成白色,用分光光度计分别在665 nm、649 nm 和 470 nm 处测定吸光度(A)值,按下列公式计算叶绿素和类胡萝卜素的含量[9]。
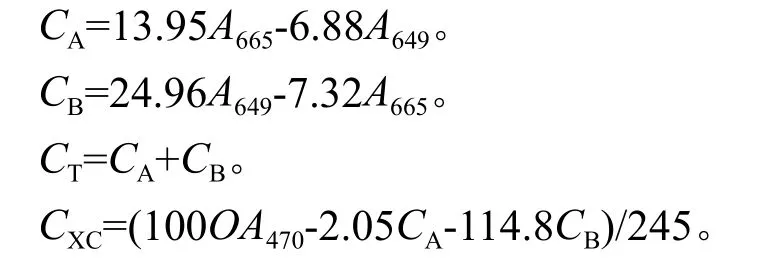
叶绿素含量(mg·g-1)=(色素浓度×提取液体积只稀释倍数)/样品鲜重,稀释倍数为3,上述测定重复3次,求其平均值。
2.4.3 光合参数测定
在8月中旬晴朗天气下,利用Li-6400便携式光合测定仪(型号为:Ci2AS)对银杏幼苗的光合生理指标日进程进行测定,主要包括净光合速率、蒸腾速率、气孔导度、胞间CO2浓度、光合有效辐射、大气温度、大气相对湿度、大气CO2浓度等光合参数。每处理均随机选择3株,每株重复3次。
3 结果与分析
3.1 生物遮阴对银杏幼苗生长的影响
生长是生理代谢过程在其外在形态上的综合反映,植物生长不仅离不开合适的土壤、水、肥、气、热和光合作用,而且更是植物体内生理代谢对外在环境的一种反馈表现。合适的光照条件对植物生长发育有着重要的作用,生物遮阴在一定成上会影响植物的生理代谢及生长状况。胁迫会导致细胞膜通透性增大,打破细胞内离子平衡,严重时会导致酶失活,生理代谢紊乱,光合作用降低,光合产物下降,植物会表现出一定程度的受害症状,表现在植物的外部形态上主要是影响其苗高、地径、叶片颜色、生物量的积累等。目前对植物幼苗的遮阴多数使用遮阳网,而大田种植采用遮阳网有所不便,因此生物遮阴对植物生长的影响越来越受到人们的关注。
3.1.1 不同遮阴强度对苗高和地径的影响
苗高、地径是最直接反映植物生长状况且容易测定的生长指标。不同的遮阴程度对银杏幼苗的苗高和地径表现出显著差异(表1),随着遮阴程度的增大,银杏幼苗的苗高和地径均比全光照要高,遮阴程度为30%和50%的均与全光照差异显著,遮阴程度为70%的与全光照无明显差异,且遮阴程度为30%时苗高和地径最高分别为14.50 cm,4.29 mm,遮阴程度为30%、50%、70%处理的苗高分别比全光照增长了42.16%、36.57%、12.45%,地径分别比全光照增粗了2.25 mm、1.56 mm、0.95 mm。可见适当遮阴有助于植物高生长和根茎部分的增粗,但遮阴程度过大反而对银杏幼苗的高生长和地径增粗的影响不大。

表1 遮阴处理对银杏幼苗生长的影响†Table 1 Effects of shading on growth of Ginkgo biloba L. seedings
3.1.2 不同遮阴强度对主根长的影响
如表1所示,遮阴30%时主根长与全光照差异显著,且主根长达到最大值8.30,而遮阴50%、70%时主根长均与全光照差异不明显,但所有遮阴处理的主根长均高于全光照的。遮阴30%、50%、70%的处理下主根长分别比全光照增长了47.42%、10.12%、21.31%。
植物根系吸收水分、养分能力的大小在一定程度上与植物的主根长有关,对植物进行遮阴处理后,植物由于得不到充足的光照,减少了水分的蒸发,促进植物根系的生长;同时过度遮阴使幼苗光合作用不够进而不能制造足够养料满足其地下生长,从而导致根系根生长受到限制。
3.2 遮阴对银杏生物量积累及其分配的影响
3.2.1 不同遮阴强度对生物量积累的影响
植物获取能量的能力大小主要是通过生物量来体现,并且生物量对植物的生长及结构的形成有一定程度的影响。植物对外界环境的适应能力主要体现在生物量的大小。
遮阴处理对银杏幼苗生物量的积累有一定的影响(表2),不同遮阴强度对银杏幼苗总生物量积累没有显著性影响(P<0.05),银杏总生物量在遮阴30%强度下达到最大,为2.333g,比全光照增加了21.13 %,但未出现显著性差异;遮阴70%处理总生物量比全光照增加了5.20%,没有显著性差异;遮阴50%的总生物量最小,比全光照减少了 12.51 %,没有显著性差异。遮阴 30 %与50%存在显著性差异,但遮阴30%与70%、50%与70%处理间没有差异性。
不同遮阴程度对银杏叶生物量未达到显著性差异(P<0.05)(表2),随遮阴程度增加叶生物量先增加后减少,遮阴30%的叶生物量最大,为1.069 g,比全光照增加了4.70%,而遮阴50%、70%的叶生物量分别比全光照减少了13.52%、6.46%。适度遮阴对叶的比重有一定的增加,从而使苗木质量变得更好,但遮阴过度反而会有抑制作用。
茎生物量在不同遮阴程度下呈显著差异(表2)。茎生物量随遮阴程度增加呈先增加后降低的趋势,遮阴30%时的生物量最大,为0.766g,比全光照增加了12.32%。遮阴50%、70%处理下的茎生物量分别比全光照减少了34.75%、17.45%,遮阴30%和50%时与全光照均存在显著差异性,而遮阴70%时与全光照未出现差异性。
根生物量在不同遮阴程度下出现了显著性差异(表2)。遮阴程度为30%的根生物最大,为0.546g,比全光照增加了51.67%;遮阴70%次之,比全光照增加了9.44%,50%分别比全光照降低了2.83%。除遮阴30%与全光照处理间存在差异性外,其它遮阴处理间及与全光照间均没有显著性差异。
植物为了在不同的光照条件下生长,会通过改变其叶片结构来适应该环境。不同遮阴程度比叶面积之间的差异达显著水平(表2),随着遮阴程度的增加,比叶面积呈现上升的趋势。遮阴程度50%、70%处理下比叶面积与全光照的均差异显著,遮阴程度30%、50%、70%处理下比叶面积分别是全光照处理下的 1.02、1.10、1.13倍。

表2 遮阴处理对银杏幼苗生物量积累和分配的影响Table 2 Effects of shading on biomass accumulation and allocation of Ginkgo biloba L.seedlings
在遮阴条件下,植物为了合成更多营养物质来满足其生长需要,会通过调整叶面积获得更多的光照来进行光合作用,这也是植物在逆境中生长时一种形态学的调节,通常比叶面积(SLA)会因光照的减弱而增大,SLA 的变化可能是植物维持最优捕获光能的一种自我平衡机制,因为在同样的干物质下具有更大的叶面积可以补偿光合有效辐射的降低[10],而全光照下则会减小叶面积来减小强光辐射造成的伤害,同时也能有效地降低水分蒸发。
3.2.2 不同遮阴强度对生物量分配的影响
在遮阴条件下,植物在受到光照限制条件下的生存能力通常表现在其地上部分与地下部分生物量的分配比例上。如表2 所示,随着遮阴程度的增加,银杏幼苗叶生物量比和根生物量比呈上升趋势,而茎生物量比呈下降趋势,导致根冠比呈先下降后上升的趋势。不同遮阴程度下的茎生物量比与全光照的均没有显著性差异,叶生物量比在遮阴为50%时达到最大值0.568,与全光照形成了显著性差异,而30%、70%处理均与全光照处理的无明显差异。遮阴50%、70%处理下根的生物量比均与全光照未形成显著性差异。遮阴30%的根生物量比与全光照呈显著性差异,且与全光照相比有所下降,遮阴30%的根生物量比值最小,为0.168,除遮阴30%处理根冠比与全光照处理存在显著性差异外,各遮阴处理与全光照处理均未达到显著性差异。
可见,随遮阴强度的增加,银杏地下部分的生物量呈增加趋势,而地上部分的生物量分配有所减少,由于地上部分受到环境条件的限制从而导致生物量分配方式向根部方向倾斜,造成根系健壮生长以便获得土壤养分来供给地上部分。
3.3 遮阴对银杏叶片叶绿素含量的影响
植物光合作用能力的大小与其叶片中叶绿素(Chl)含量多少密切相关。而叶片中的叶绿素含量的多少又会随着光照条件的变化而变化,从而使植物更好的适应在不同光照条件下生长。进行光合作用的主要色素是叶绿素a和叶绿素b,叶绿素a的功能主要是将汇聚的光能转变为化学能进行光化学反应,而叶绿素b则主要是收集光能,叶绿素a/b比值可以判定逆境对两种色素的影响程度。类胡萝卜素(Car)也有收集和传递光能的作用,除此之外,还有防护叶绿素免受多余光照伤害的功能[8]。
不同程度遮阴对银杏叶片中叶绿素含量的影响见表3,由方差分析可知,在遮阴条件下叶绿素a含量均比全光照的有所增加,遮阴30%时叶绿素a含量最高,为 1.677 mg·g-1,是全光照的1.90倍,且与全光照存在显著差异。不同处理间均存在显著差异性,但70%处理与全光照下出现显著性差异;遮阴50%、70%叶绿素a含量分别是全光照的1.30、1.13倍。

表3 遮阴对银杏叶片叶绿素含量的影响Table 3 Effects of shading on chlorophyll content of Ginkgo biloba L.seedlings
遮阴条件下叶绿素b含量均比全光照下有所增加(表3),遮阴30%、70%处理下叶绿素b含量均与全光照存在显著差异,但二者之间没有差异性,遮阴程度为30%、50%、70%的叶绿素b含量分别是全光照的1.46、1.11、1.47倍。
遮阴下叶绿素(a+b)含量均有所增加,遮阴程度为30%时的含量最大,遮阴30%、50%处理的叶绿素(a+b)含量均与全光照存在显著差异,遮阴处理下的叶绿素(a+b)的含量分别是全光照处理的1.78、1.25、1.22倍。遮阴30%、70%处理下叶绿素 a/b 的值与全光照处理呈现显著差异性,随着遮阴程度的增加叶绿素a/b表现为先上升后下降的趋势,遮阴30%处理下叶绿素 a/b 的值最大为3.319,是全光照下的1.31倍,而遮阴70%条件下含量却减少。
银杏叶片中类胡萝卜素(Car)含量在遮阴条件下均比全光照下要增加(表1~表3),遮阴30%时含量最高0.387 mg·g-1,与全光照处理形成了显著性差异;而50%、70%处理未出现差异性。遮阴30%、50%、70%处理下类胡萝卜素(Car)含量分别是全光照的1.38、1.20、1.18 倍。遮阴30%处理下类胡萝卜素/(叶绿素a/b)比值最小,且与处理间及全光照下均出现显著性差异,除遮阴30%处理与全光照间存在差异外,其它遮阴处理与全光照均没有显著差异性,遮阴30%、50%、70%处理下类胡萝卜素/(叶绿素a/b)的比值分别是全光照的0.78、0.96、0.97 倍。
3.4 遮阴对银杏光合生理变化的影响
3.4.1 气孔导度和胞间CO2浓度变化
气孔作为陆生植物叶片上一个重要的结构,它是植物与外界进行水分和CO2等气体交换的重要门户。气孔的开闭会对植物叶片的光合和蒸腾产生影响,而气孔导度则是反映植物气孔行为最重要的生理指标。
气孔导度(Gs)随着遮阴强度的增加而减小(见表4),Gs在遮阴为70%时的值最小,且与全光照有显著差异,各处理间差异不明显。遮阴30%、50%、70%处理下Gs的值分别全光照的0.75、0.71、0.57倍。
随遮阴强度为30%的胞间CO2浓度(Ci)的值最小为330,与全光照有显著差异,且是全光照的0.87倍,而遮阴为50%、70%处理下胞间CO2浓度(Ci)均与全光照无明显差异。但都比全光照的值要小。

表4 遮阴对银杏叶片光合作用的影响Table 4 Effects of shading on photosynthesis of Ginkgo biloba L. seedlings
3.4.2 净光合速率和蒸腾速率的变化
净光合速率(Pn)是一定时间内,植物同化的总光合产物与呼吸消耗的光合产物之差,是研究植物光合作用状况的最重要指标[11]。随着遮阴程度增加,净光合速率逐渐减小,分别是全光照的92.96%、81.69%和54.93%。张往祥等经过2年银杏遮阴试验研究发现,净光合速率随遮阴强度加大而下降的幅度远小于光合有效辐射的下降幅度,遮阴有利于减轻甚至避免银杏光合作用的“午休”现象。不同遮阴处理下蒸腾速率Tr与全光照未出现差异性,这可能与银杏幼苗期具有一定耐阴性有关[2]。
4 结 论
(1)一般来说,合成叶绿素的适应温度为30 ℃[12],而适合银杏叶片合成叶绿素的最佳温度为26 ℃左右[13]。由于夏季温度过高,不利于叶绿素的合成,适当地遮阴有利于增加叶片中叶绿素的含量,从而光合作用变强,产生更多的光合产物供给苗木生长,使苗木长得更加健壮。
(2)遮阴有利于降低叶片的温度,防止叶片在夏季高温时被强光灼伤,同时也降低植物的蒸腾作用,减少叶片中的水分散失,在这个意义上,适当地对植物进行遮阴更有利于提高植物光合作用。
(3)银杏幼苗具有一定的耐荫性,在幼苗生长期间,适当地采取遮阴措施引起植物叶面积的增加,有利于银杏在低光照强度下能够获得更多光照,但遮阴程度应在有限范围内[14]。本次实验结果为银杏叶用林的栽培和管理技术提供了一定的理论基础,因此叶用林的栽植密度不宜过大。
