伊犁河谷不同造林模式土壤养分与酶活性的关系
2018-07-26桑巴叶朱玉伟陈启民王永红褚奋飞
桑巴叶, 朱玉伟, 陈启民, 王永红, 褚奋飞
(新疆林科院 造林治沙研究所, 新疆 乌鲁木齐 830063)
森林是陆地生态系统的重要组成部分,是地球上最为宝贵的自然资源,一个独特的人类和多种生物赖以生存和发展的生态系统[1]。在森林生态系统中,土壤与植被密不可分,构成一个复杂的功能体系。土壤为森林植物及土壤生物提供生长定居的机械支持和物质基础。森林植物在生长过程中所需养分极大部分通过植物本身新陈代谢过程中或自然条件调节所形成的凋落物、动物残体在土壤中经过分解成小分子化合物,供给森林植物生命所需[2-3]。不同的森林类型提供的物质基础的差异,影响微生物种群数量变化的同时亦影响土壤物理化学性质和养分数量[4-5],造成森林土壤生产力的差异。
土壤酶主要指土壤中的聚积酶,包括游离酶、胞内酶和胞外酶,来源于土壤微生物的活动、植物根系分泌物和动植物残体腐解过程中释放的酶。土壤酶参与土壤中的一切生物化学过程,催化了土壤中众多生化反应[6-7]。由于土壤酶活性与土壤理化性质、土壤生物数量和生物多样性等密切相关,因此可以作为土壤质量的生物活性指标[8]。研究不同林分类型对土壤酶活性的影响将对林区生态学与土壤学的研究起到积极推动作用,因此土壤酶活性已然成为目前森林生态系统研究中必不可少的内容。
伊犁河谷位于新疆西天山地区,是新疆及天山气候最湿润、降水最丰沛、植被土壤发育最良好的地区,森林覆盖率为7.9%,林、草植被覆盖率高达67.7%,被喻为新疆干旱区的一块“湿岛”。对伊犁河谷土壤的研究,将会更加充分地理解植被状况对土壤质量和环境的潜在影响,最终为森林土壤质量的评价提供依据。
近年来,国内外学者从不同角度对土壤类型、土壤理化性质、土壤酶活性等方面进行了广泛研究。对于森林土壤养分的研究已有大量的研究报道,研究内容涉及养分时间动态变化、土壤养分储量、土壤养分的空间动态变化等方面[9]。关于林分类型对土壤理化性质的影响,已经取得了重要的研究成果[10]。土壤酶作为土壤质量的生物活性指标,自20 世纪 80 年代以来已经被大量研究[11]。目前广大学者对土壤生态因子和林分类型与土壤酶之间的相互关系十分关注,并做了相应的研究与报道[12]。关于伊犁河谷森林土壤养分与酶活性方面研究仅局限于单一林分[13]、单一酶活性[14]或养分[13]、不同土地利用方式下土壤养分和酶活性研究[15]等方面,其它相关研究报道极少。伊犁河谷造林模式多样,存在着长期选择和演替的规律性变化,故对不同造林模式下土壤养分和酶活性特征的研究十分必要。因此,本研究拟以伊犁河谷地区为研究区域,选择当地典型的5种造林模式为研究对象,分析土壤养分和土壤酶的变化状况。通过研究不同造林模式下土壤养分和酶活性变化规律,以期为伊犁河谷地区造林树种的搭配和布局提供科学依据。
1 材料与方法
1.1 研究区概况
伊犁河谷位于中国天山山脉西部,准噶尔盆地西南缘,属南天山、北天山分隔而成的由断陷盆地与山间河谷间布的中亚内陆河流域。地理位置为东经80°09′42″—84°56′50″,北纬42°14′16″—44°53′30″。三面环山,流域地跨中温带大陆性气候和高山气候类型,气候温和湿润,昼夜温差大。年均降水量200~800 mm,降水随高程上升增多明显。平均海拔高度为477~1 701 m,气候温和湿润,降水量充足,是中国唯一的温带大陆性气候地区,部分地区属于高山气候。伊犁河谷地域辽阔,资源丰富,素有“塞外江南”的美称。土壤类型多样,包括潮土、灌耕土、草甸土、黑钙土、栗钙土、灰钙土、沼泽土等。
1.2 研究方法
1.2.1 造林模式的选择 选取伊犁河谷5种典型生态造林模式,即杨树用材林、沙枣防护林、杨树防护林、榆树防护林和林草间作用材林。均为成熟林。不同造林模式样地基本情况见表1。

表1 不同造林模式样地基本情况
1.2.2 土壤样品的采集 在对研究区实际调查的基础上,在5种造林模式中各设置3个标准样地,样地规格为林带宽×50 m,按“S”型随机采取5个样点的土样,在每个样点取0—20和20—40 cm土层土样,将相同土层的样品混合,去除石块、根系和土壤动物等,经实验室风干、过筛后贮存备用。
1.2.3 测定指标及测定方法 ①土壤化学性质测定[16]:土壤全氮采用半微量凯氏定氮法测定;土壤全磷采用氢氧化钠熔融、钼锑钪比色法测定;土壤全钾采用氢氧化钠碱熔、火焰光度计法测定;土壤水解氮采用碱解扩散法(样品正在分析中);速效磷用钼锑抗比色法,速效钾用醋酸铵浸提—火焰光度计法测定;土壤有机质采用重铬酸钾容量法—外加热法。②土壤酶活性测定[17]。土壤过氧化氢酶和脲酶采用《土壤酶及其研究法》进行测定。
1.3 统计分析
试验数据采用Excel 2007和SPSS 19.0 软件进行统计分析。
2 结果与分析
2.1 土壤养分特征
树种影响着水分、土壤、植物和微生物之间的相互作用,各模式中的树种不同,土壤养分就会呈现出不同的差异。由表2可知,0—20 cm土层林草间作用材林的有机质含量最高,次之为杨树防护林,均与其他模式之间差异显著(p<0.05),杨树用材林、沙枣防护林和榆树防护林之间差异不显著(p>0.05)。林草间作用材林和沙枣防护林20—40 cm土层有机质含量分别为最高和最低,含量最低值仅为最高值的24.82%。各模式土壤有机质平均含量高低顺序为:林草间作用材林>杨树防护林>榆树防护林>杨树用材林>沙枣防护林。除榆树防护林外,其余模式均随土层深度增加,土壤有机质含量下降,其中杨树防护林降幅最大,杨树用材林最小。
土壤全氮在2个土层之间表现出了相似的规律,林草间作用材林全氮含量最高并与其他模式之间差异显著,沙枣防护林最低,林草间作用材林全氮含量为沙枣防护林含量的3.46倍。各模式土壤全氮均值:林草间作用材林>榆树防护林>杨树防护林>杨树用材林>沙枣防护林。碱解氮0—20 cm土层榆树防护林碱解氮含量最高,林草间作用材林次之,两者之间没有显著差异,但均与其他模式之间有显著差异,2个土层碱解氮高低顺序一致,均为榆树防护林>林草间作用材林>杨树用材林>沙枣防护林>杨树防护林。
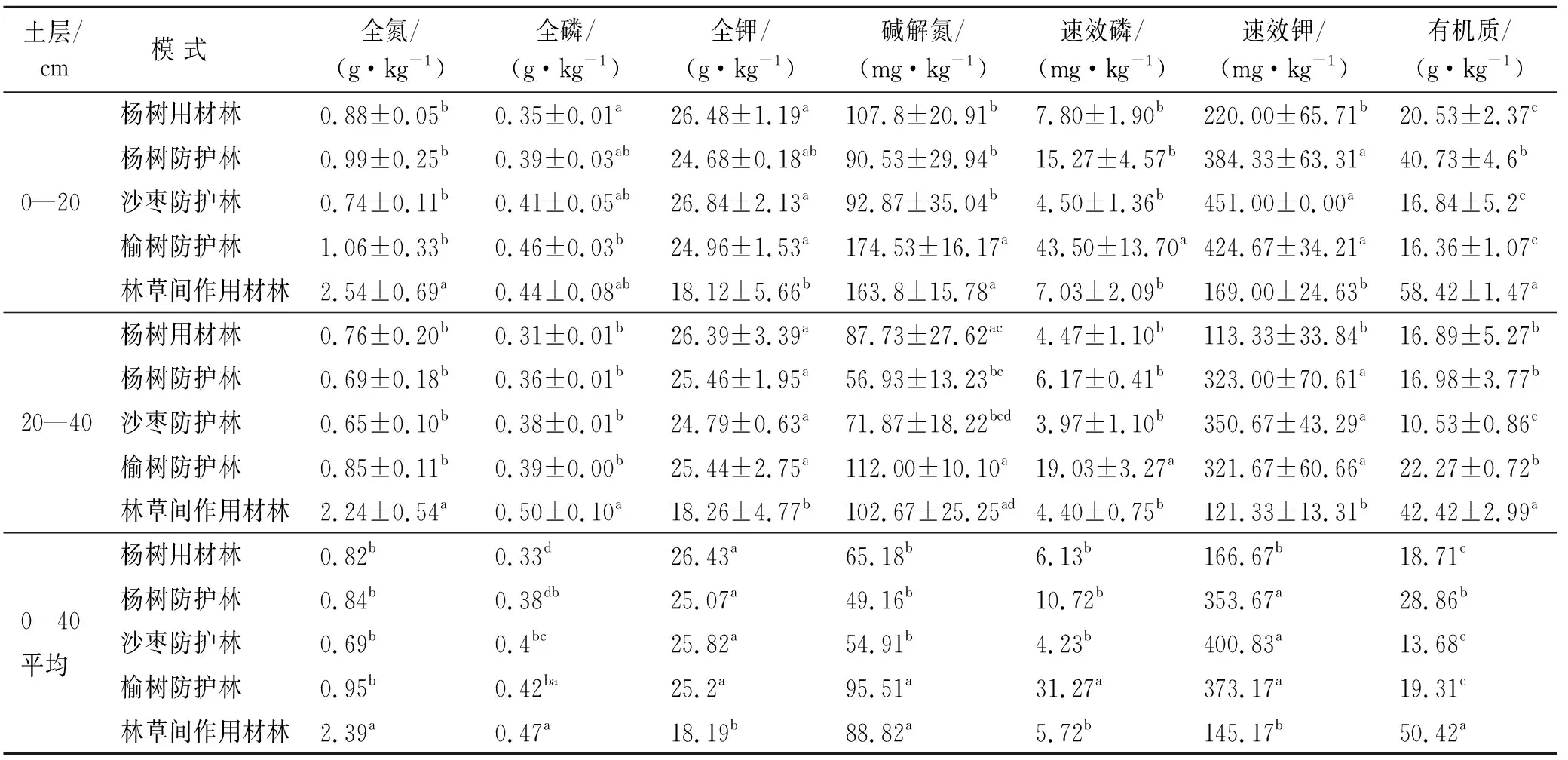
表2 伊犁河谷不同造林模式土壤化学性质
由表2可以看出各模式林分土壤全磷含量总体较低,范围在0.31~0.50 g/kg,各模式之间相关不大。0—20 cm土层榆树防护林全磷含量最高,最底为杨树用材林,20—40 cm土层全磷含量最高则为林草间作用材林。各模式林分土壤全磷均值表现为林草间作用材林〉榆树防护林〉沙枣防护林〉杨树防护林〉杨树用材林。各模式间土壤速效磷含量相差较大,2个层次的含量在3.97~43.5 mg/kg之间,速效磷含量最高值为最低值的11倍。0—20 cm土层含量最高的榆树防护林为最低的沙枣防护林的9.67倍,20—40 cm土层相同情况,最高为最低的4.79倍。随土壤深度增加,有效磷含量急剧降低,其中杨树防护林和榆树防护林降幅最大。榆树防护林与其他模式之间差异显著,其他各模式之间差异不显著。有效磷均值高低顺序为:榆树防护林>杨树防护林>杨树用材林>林草间作用材林>沙枣防护林。
5种模式2个层次土壤全钾的含量为18.12~26.84 g/kg,速效钾含量为121.33~384.33 mg/kg,均属于较高水平。5种模式的2个层次之间全钾含量变化幅度小。在0—20 cm土层沙枣防护林含量最高,20—40 cm则为杨树用材林。各林分土壤全钾均值排序:杨树用材林>沙枣防护林>榆树防护林>杨树防护林>林草间作用材林。沙枣防护林速效钾含量在2个土层中最高。0—20 cm土层中林草间作用材林速效钾含量最低,仅为含量最高的沙枣防护林的37.47%。20—40 cm最低的林草间作用材林仅为含量最高的沙枣防护林的32.29%。沙枣防护林、榆树防护林和杨树防护林2个土层中的速效钾含量均属很高水平,与其他2个模式之间差异显著。各模式林分土壤速效钾均值表现为:沙枣防护林>榆树防护林>杨树防护林>杨树用材林>林草间作用材林。
2.2 土壤酶活性特征
图1为不同造林模式下土壤酶活性差异。图1表明,不同造林模式土壤土层酶活性的变化趋势基本一致,0—20 cm土层酶活性均高于20—40 cm土壤酶活性。

注:不同字母表示同一土层同一酶活性在不同造林模式中差异显著(p<0.05)。
脲酶参与有机态氮素的分解作用, 专门水解有机氮转化过程的中间产物尿素, 使其转化成为氨被林木利用, 其活性可以用来表示土壤氮素供应状况。杨树防护林0—20 cm脲酶活性最高,为2.32 mg/g,榆树防护林20—40 cm脲酶活性最高,为1.61 mg/g。沙枣防护林脲酶活性最低,0—20 cm土层为0.86 mg/g,20—40 cm仅为0.40 mg/g。不同造林模式0—40 cm 土层土壤脲酶活性高低排序为:榆树防护林>杨树防护林>林草间作用材林>杨树用材林>沙枣防护林。
过氧化氢酶是土壤中一种重要的氧化还原酶,参与土壤物质和能量转化过程,具有分解土壤中对植物有害的过氧化氢的作用。研究表明,过氧化氢酶活性能够反映土壤的呼吸强度[18]。不同造林模式0—20土层土壤过氧化氢酶活性变化范围在0.20~1.57 mg/g,20—40 cm土层在0.18~1.50 mg/g。杨树防护林酶活性为最高,比活性最小的林草间作用材林高8.1倍。不同林分土壤过氧化氢酶的排列顺序为:杨树防护林>沙枣防护林>杨树用材林>榆树防护林>林草间作用材林。
2.3 土壤酶活性与土壤养分的相关性分析
研究表明[19-22],土壤酶活性与土壤养分互相作用,土壤养分含量的高低制约着土壤酶活性的大小,反之,土壤酶活性的反馈作用使土壤养分含量发生变化,二者之间存在着非常密切的相关关系。为此,我们对土壤酶活性与土壤养分之间进行相关分析。研究结果表明(表3),在表土层,脲酶和除全钾外其他呈正相关,其中和速效磷呈显著正相关(p<0.05);过氧化氢酶和全钾、速效钾呈显著正相关(p<0.05),和其他养分指标呈负相关,其中和全氮、碱解氮有关系最为密切,均达到极显著负相关(p<0.01),和有机质呈显著负相关(p<0.05)。
在底土层,脲酶和全钾呈负相关,与其他养分指标呈正相关,其中和速效磷、有机质呈显著正相关(p<0.05);过氧化氢酶和全氮、有机质呈极显著负相关(p<0.01),和全磷、碱解氮呈显著负相关(p<0.05),和全钾、速效钾呈显著正相关(p<0.05)。
在整个土层中,脲酶和全钾呈负相关,和其他养分指标呈正相关;过氧化氢酶和全钾、速效钾呈正相关,和其他养分指标负相关。因此,不同类型的土壤酶活性与土壤养分之间存在着相关关系,故土壤酶活性高低情况可作为评价土壤肥力的指标。

表3 伊犁河谷土壤酶活性与土壤养分的相关分析
3 结 论
(1) 在同一造林模式林分内,土壤养分含量主要分布在0—20,20—40 cm土层则较低。不同造林模式土壤肥力状况有明显的差异,杨树用材林的全钾含量最高,杨树防护林的有效钾含量较高,沙枣防护林土壤全钾和有效钾含量最高,榆树防护林土壤碱解氮和有效磷含量最高,全钾和有效钾处于较高水平,林草间作用材林土壤全氮、全磷和有机质含量最高,碱解氮也处于较高水平。凋落物是森林生态系统的重要物质库,通过植被枯落物的形式能不断地给予土壤养分,使得土壤表层养分含量显著增加,进而达到互利共赢。
(2) 土壤酶活性是森林生态系统的重要组成成分,对土壤养分的流动与转化起着重要作用,可以客观地反映土壤质量状况。土壤酶活性不仅与土壤理化性质有关,而且与林分类型也密切相关。在土壤空间上,脲酶和过氧化氢酶活性随土层的增而减小。在整个土层中土壤脲酶活性排序为:榆树防护林>杨树防护林>林草间作用材林>杨树用材林>沙枣防护林;过氧化氢酶活性排序为:杨树防护林>沙枣防护林>杨树用材林>榆树防护林>林草间作用材林。
(3) 土壤养分与土壤酶活性之间存在一定的相关性。在整个土层中,脲酶和全钾呈负相关,和其他养分指标呈正相关,其中和全磷显著正相关(p<0.05),和碱解氮、速效磷、有机质极显著正相关(p<0.01);过氧化氢酶和全钾、速效钾呈极显著正相关,和全氮、全磷、碱解氮、有机质呈极显著负相关。本研究结果同李传荣等[23]的研究结果一致,但与其他多数文献结果不同,研究发现土壤中过氧化氢酶、脲酶活性与土壤主要养分因子间均存在极显著或显著的正相关关系[19-22]。这种存在不一致情况,需在今后的研究中进一步考证。
