不同栽植代数杉木林养分吸收、积累和利用效率的比较
2018-07-18周玉泉康文星陈日升田大伦项文化
周玉泉, 康文星,2,3,*, 陈日升 , 田大伦,2,3,项文化,3
1 中南林业科技大学,长沙 410004 2 南方林业生态应用技术国家工程实验室,长沙 410004 3 国家野外科学观测研究站,会同 418307
19世纪中期,当Ebermayer测定了阔叶林和针叶林的养分含量[1],揭开了森林养分动态研究序幕,20世纪60年代后,Cole[2]、Duvigneaud[3]、Bormann[4]、Tsutuumi[5]、Bazilevich[6]等国外学者在森林养分的研究方法、测定技术、养分循环的分类等方面做出了重要贡献。
20世纪50年代侯学煜等已分析了中国150种植物化学成份[7]。20世纪70年代后,潘维俦[8]、冯宗炜[9]等分别开启了杉木(Cunninghamialanceolata(lamb) Hook)人工林养分循环的研究。随后我国进入一个关于森林养分动态研究的快速发展阶段,尤其近10多年许多学者对不同类型的森林,尤其是人工林[10- 16]养分循环展开大量研究。这些研究成果都具有重要的实践和理论意义。以上国内大部分研究主要集中在林分某一生长阶段的养分积累与分配的静态特征上。虽然也有学者探讨了不同生长阶段林分养分积累与分配的动态变化[17- 20]。但大都用“空间换时间”的研究方法。其研究结果可能偏移实际。
19世纪初就已发现人工林连栽后会出现生产力下降和地力衰退的“二代效应”问题[21]。为探索杉木连栽地力衰退的原因,有学者从林分养分动态出发采用时空转换法对不同代数杉木林养分循环进行比较[22-23]。也有学者对第一代杉木林采伐后紧接营造第二代杉木林的两代杉木林的养分利用效率进行分析[24],由于估算不同生长阶段林分净生产力和每年吸收的养分量的方法不完全准确,其研究结果难免会有疵瑕。本研究用会同第二代与第一代连栽的两代林40多年的生物量和养分测定数据,采用新的研究方法(在研究方法中说明),分析两代杉木林在不同生长阶段的养分吸收、积累以及利用的差异。从两代杉木林的养分动态变化上,探讨人工林连栽后会出现生产力下降和地力衰退的原因。为人工林的持续经营和管理提供科学依据。
1 材料与方法
1.1 研究区概况
本研究在湖南会同杉木林生态系统研究站(属于国家野外科学—观测站)第3试验小集水区进行。地理位置 26°50′N,109°45′E,面积1.984 hm2,海拔270—350 m,坡向西北,坡度15—25°。研究区属于中亚热带季风湿润气候区,年平均气温16.9℃,年降雨量1100—1400 mm,年均相对湿度80%以上。土壤母岩为震旦纪板溪系灰绿色板岩,风化程度较深,土层厚度80 cm以上,土壤为中有机质厚层黄壤。
1962年第3小集水区在原为残存的阔叶次生林地营造杉木人工林,其杉木林于1987年冬皆伐了,并于1988年春在采伐迹地上营造的第二代杉木林。两代林的整地造林方式相同(炼山、全垦挖穴),造林密度也相近(约3000株/hm2),而且都是杉木栽上后前3 a的春秋季节各抚育1次,以后任其自然生长。
1.2 样品采集
第3小集水区第一代杉木人工林7 年生时,设立了4块固定标准地(样地面积667 m2)测定林分生物量,以后在第一代杉木人工林11、16、20年生和25年生时,都在4块固定标准地测定林分生物量。第二代杉木人工林营造后,同样设立了4块固定标准地,并于第二代林7、11、16、20年生和25年生时,在4块固定标准地测定林分生物量。每次测定生物量的同时采集养分待测样品。为了保证样品具有代表性,在每一次采集样品的伐倒的标准木上把它分成树干基部、中下、中、中上和上部5个区段,每一区段釆集相同重量的树干,再把釆集的树干混合一起组合成树干样品;树皮取样采用树干相同的方法。枝和叶都分成当年生、1 年生、2 年生、多年生分别取样。根分成根头、大根(1 cm≤根径)、粗根(0.2 cm≤根径< 1 cm)、细根(根径<0.2 cm)分别取样。
1.3 养分测定
将待测的杉木各器管组织样本,取适量样品置于85℃恒温干燥箱内烘干至恒重,称干重,磨碎,过60目筛后按《土壤农业化学分析方法》[25]配置待测溶液。植物中的全N用半微量凯氏法,全P 用分光光度计测定;全K、全Ca、全Mg均用原子吸收分光光度计测定。
1.4 不同生长阶段养分积累现存量计算
林分某一林龄时养分积累现存量用下式计算:
M1=W1i×L1i
(1)
式中,M1表示林分某一林龄时养分积累现存量(kg/hm2);W1i表示林分某一林龄时杉木某器官现存生物量(t/hm2);L1i表示林分某一林龄时杉木某器官的养分浓度(g/kg);i=1,2,…,5(干、皮、枝、叶、根) 。
1.5 不同生长阶段养分吸收量计算
以往林分某生长阶段的养分吸收量都采用“吸收=存留+归还”的计算方法[3]。这只有在某生长阶段林分归还的养分,完全是这生长阶段生产的物质的枯死物中的养分时才能成立。实际上林分某生长阶段的枯死物并不完全是这个生长阶段生产的物质中枯死的(尤其是常绿林分),有一部分可能是前一生长阶段生产的物质存留下来在本生长阶段才枯死物质。因此,要准确估算某生长阶段的养分吸收量,首先要准确估算某生长阶段林分生产的生物产量。
1.5.1不同生长阶段的生产的生物产量估算
一般认为树木的干、皮在树木的生长过程中凋落甚少,随林龄增长其生物量不断积累,可以用两个不同生长阶段的这些器官现存量之差求得某生长阶段的生物产量,但枝、叶在树木的生长过程中会不断凋落,所以,要准确估算某生长阶段林分生产的生物产量,就有确定叶和枝从生长出来到枯死的整个生长期。在此基础上再估算出测定的该生长阶段林分现存枝叶生物量中有多少是前一生长阶段留下的,而且还要估算出该生长阶段内生长的枝叶在这生长阶段的枯死量。那么某生长阶段生产的生物产量(W)等于此次测定树干、皮、根的现存生物量(W2i)减去前一生长阶段的树干、皮、根现存生物量(W1i)(根系也存在更新代谢过程,但准确测定根系的周转周期,在技术上还存在很大难度,因此,本研究将根与干材和树皮同等对待,其估算出的生产量要低于实际生产量),加上此次测定的现存枝叶生物量(W3j)减去此次测定的现存枝叶生物量中属于前一生长阶段生长留下的枝叶量(W4j)的差,再加上该生长阶段生长的枝叶的枯死量(W5j)。其公式为:
W= (W2i-W1i)+(W3j-W4j)+W5j
(2)
式中,i=1,2,3(干、皮、根);j=1,2(枝、叶)。本研究中杉木叶的生长期采用刘爱琴等[22]研究成果定为5年,根据我们在会同对杉木生长的长期观测,把杉木枝生长期定为10年。
1.5.2不同生长阶段的养分吸收量计算
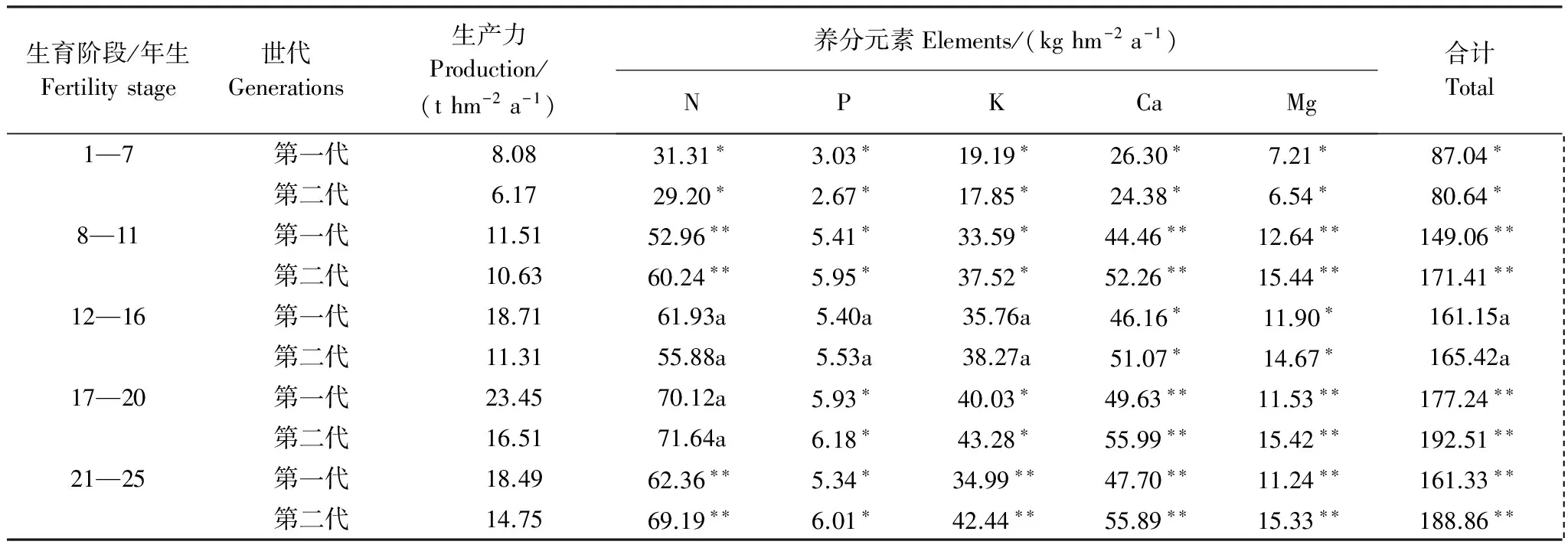
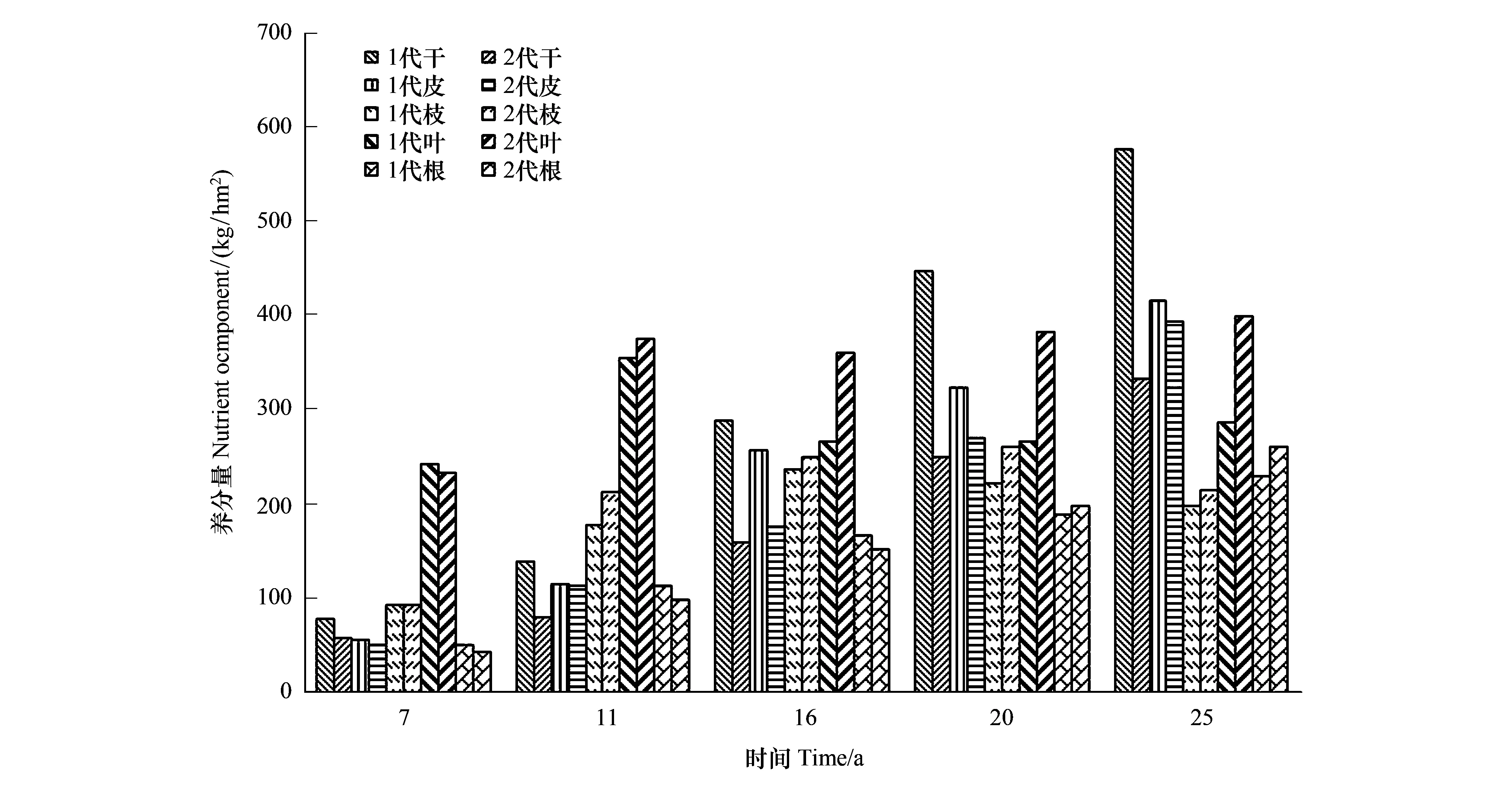
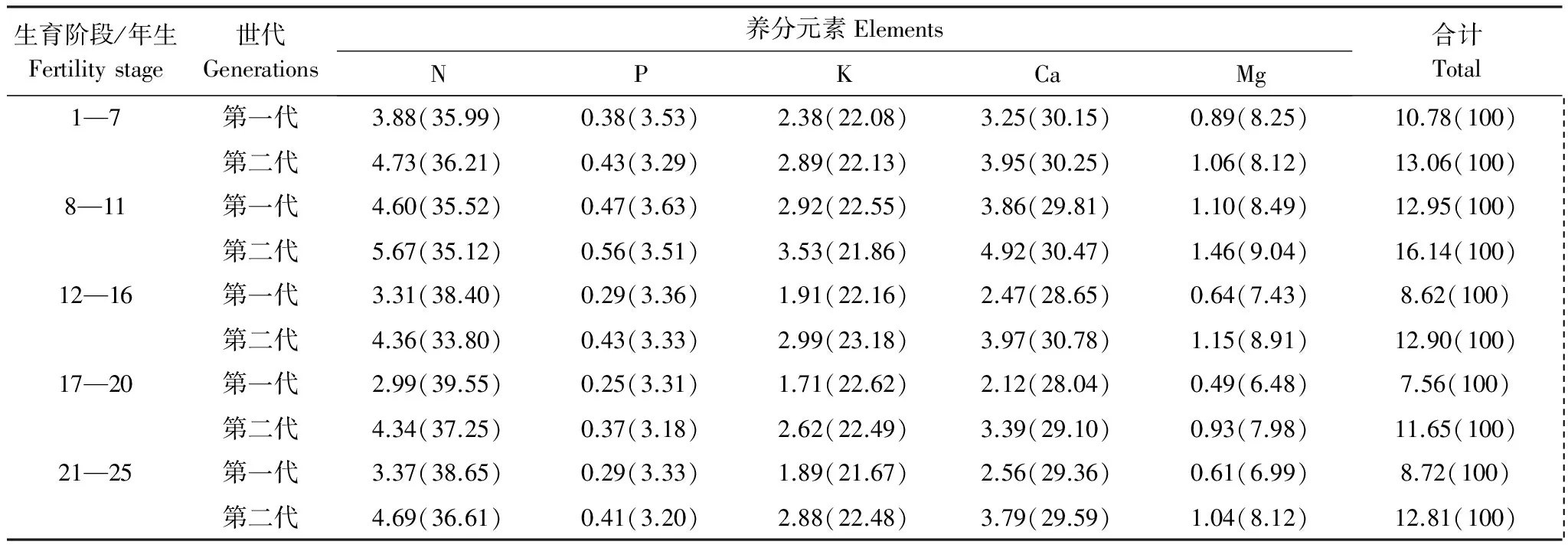
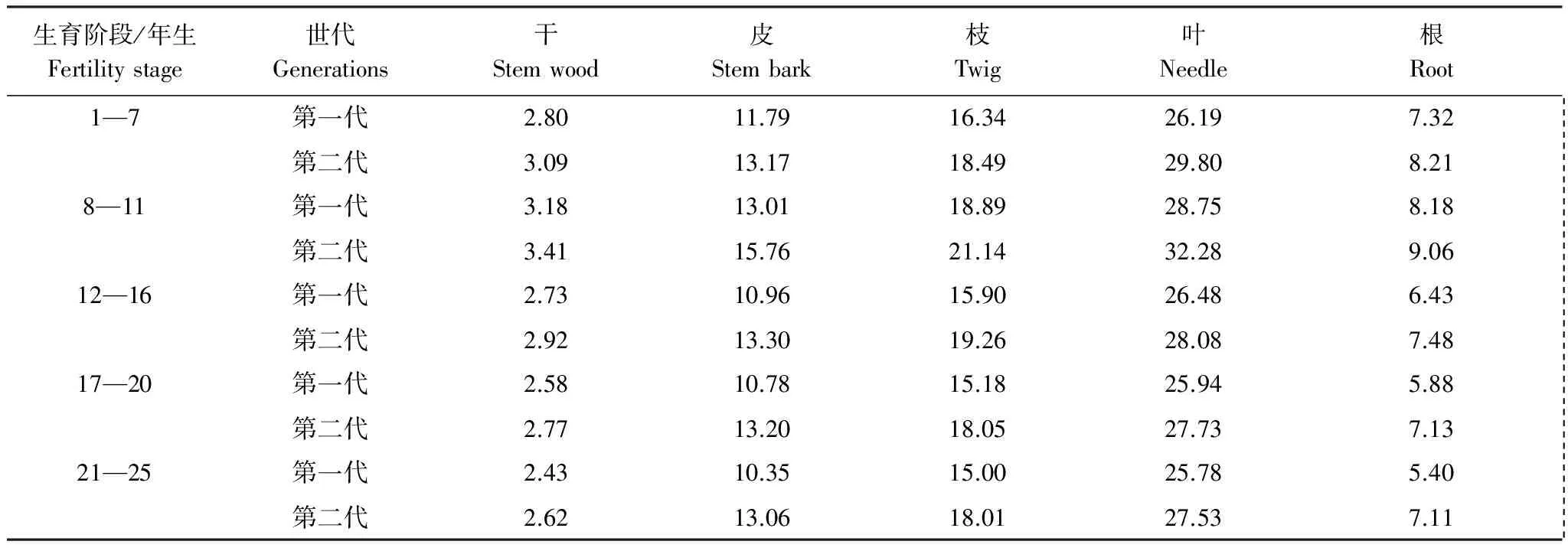
“吸收=存留+归还”也只有在某生长阶段杉木各器官养分浓度与前一生长阶段测定的杉木各器官养分浓度相等时才正确。如果某生长阶段测定的杉木各器官养分浓度为L1i,前一生长阶段测定的浓度为L2i。若L1i M3=W3i×L1i-W2i×(L2i-L1i) (3) 当L1i>L2i时,则有 M3=W3i×L1i+W2i×(L1i-L2i) (4) 式中,M3表示吸收养分量(kg/hm2);W3i表示某生长阶段生产量(t/hm2);W2i表示前一生长阶段的现存量(t/hm2);i=1,2,…,5(干、皮、枝、叶、根)。 养分利用效率的高低用每生产1 t干物质需要的养分量来表征,其需要的养分量越低,养分利用效率越高,反之则然。其计算公式为: N=M/W (5) 式中:N表示养分的利用效率(kg/t);M表示某生长阶段林分吸收养分量(kg);W表示某生长阶段林分的生产量(t)。 所有数据经Excel 软件处理,用SPSS 20.0软件进行统计分析,采用单因素方差分析(one-way ANOVA)进行不同代数,不同生长阶段,不同器官养分浓度、养分积累和分配的显著性检验。 图1 两代杉木林不同生长阶段的养分元素浓度Fig.1 Concentrations of nutrient elements in different growth stages of two generation Chinese fir forests 图2表明,同一林龄时,第二代的杉木各器官体内养分浓度也分别高于第一代。其中,树干、皮、枝、叶和根的养分浓度分别比第一代高6.86%—8.79%,11.71%—20.60%,13.17%—21.06%,10.82%—15.25%,9.76%—17.78%。经差异显著性检验表明,同一林龄时,除两代杉木的树干养分浓度之间差异显著(P<0.05)外。其余同一器官养分浓度之间差异极显著(P<0.01)。 图2 两代杉木林不同生长阶段各器官的养分浓度Fig.2 Nutrient concentrations of organs in different growth stages of two generations Chinese Fir Plantation 通过对两代林的杉木器官养分浓度随林龄增加的变化分析发现,7—11年生时养分浓度增加的速率第二代杉木是第一代的1.4倍,12—16、17—20、21—25年生时养分浓度下降的速率第二代杉木分别只有第一代的81.41%,40.56%和61.35%。 从表1中发现,1—7年生,第一代的年均吸收养分量比第二代多7.94%(7.01 kg/hm2),8—11,12—16,17—18、21—25年生,又分别比第二代少13.04%(22.40 kg/hm2),2.52%(4.27 kg/hm2),7.93%(15.27 kg/hm2)和14.58%(27.53 kg/hm2)。1—7年生第一代的N、P、K、Ca、Mg各元素的年均吸收量比第二代分别多6.74%,11.88%,9.68%,7.30%,8.88%;8—11、12—16、17—20、21—25年生的N、P、K、Ca、Mg年均吸收量分别比第二代少2.27%—13.75%,2.41%—12.25%,7.02%—21.29%,10.64%—17.91%,22.15%—36.39%。从上分析看出,林分7年生以前第一代的年均吸收养分量比第二代多,7年生后第一代比第二代少。 表1 两代杉木林不同生长阶段年平均吸收的养分 本表的差异分析是两代林同一林龄同一元素吸收量的差异分析, *表示同一林龄同一元素吸收量差异显著(P<0.05),**差异极显著(P<0.01),小写字母相同表示同一林龄同一元素吸收量差异不显著(P>0.05) 从两代林养分吸收年均量随林龄增加的变化分析中看出,第一代从1年生到20年生的养分吸收量随林龄增加而增加,20年生后随林龄增大逐渐下降,年变化曲线为抛物形,抛物线顶点出现在17—20a。第二代的1—7a和17—20年生时,年均吸收养分呈上升趋势,12—16a和21—25a呈下降趋势,年变化曲线为双波峰曲线。波峰分别出现在1—7a和17—20a。第一代8—11,12—16a和17—20a年均吸收养分上升的速率分别为71.25%,8.35%和9.98%,比第二代8—11a和17—20年生时的112.56%和16.50%小许多。第二代8—11a和17—20年生年均吸收养分下降速率分别为3.50%,1.90%,比第一代20—25a下降速率8.98%又低许多。表明林分整个生长过程中对林地养分的消耗,第一代比第二代少些。 两代林同一林龄时年均吸收同一养分元素量的差异分析表明,除12—16a的两代林的N之间、P之间、K之间,吸收总养分量之间,以及17—20a的两代林的N之间的年均吸收量差异不显著外(P>0. 05),其余两代林的各营养元素的年均吸收量之间差异显著(P<0.05),或极显著(P<0. 01)(表1)。 2.3.1养分积累的比较 从表2可知,第一代7、20、25年生时的养分积累量分别比第二代多43.02 kg/hm2(9.14%),32.06 kg/hm2(2.01%),3.96 kg/ hm2(0.22%)。11、16年生时第一代的养分积累量分别比第二代少54.42 kg/hm2(6.72%),45.48 kg/hm2(3.44%)。杉木连栽并不一定导致第二代林乔木层养分积累减少。第一代的N、P元素积累量分别比第二代多6.38—51.15 kg/hm2,0.99—2.86 kg/hm2。第一代7年生时Ca、Mg元素积累量分别比第二代多11.62和3.85kg/hm2,7年生后各生长阶段分别比第二代少9.93—25.19,8.23—22.52kg/hm2。K元素积累量,7、20a和25年生时第一代比第二代分别多8.88%,2.43%和0.97%,11a和16年生又分别比第二代少4.13%和2.03%。就总体而言,第一代的N、P、K素积累量高于第二代, Ca、Mg低于第二代。 该术式既避免了脊髓后移,防止出现C5神经根麻痹等并发症[32],又避免了直接切除骨化物,大大降低了脑脊液漏、脊髓损伤等并发症的发生率。同时,ACAF对后路手术治疗OPLL的再手术翻修也取得了良好效果[33]。Sun等[34]通过对20例OPLL患者的CT资料进行研究,认为钩突关节可作为ACAF纵向截骨开槽位置的标志,但术前还应对每个病例的CT图像进行仔细的测量以制定个体化的精确手术方案。 表2 两代杉木人工林不同林龄时乔木层积累的营养元素量 括号内的数为百分数,本表的差异分析是两代林同一林龄同一元素积累量的差异分析, *表示同一林龄同一元素积累量差异显著(P<0.05),** 差异显著(P<0. 01),小写字母表示同、同一林龄间同一元素积累量差异不显著(P>0.05) 两代林同一林龄时各营养元素积累的差异分析表明,除11年生时两代林的N之间、P之间、 K之间,16年生时两代林的N之间、P之间,20 年生时两代林的K之间、Ca 之间,25 年生时两代林的K之间、P之间,养分总积累量之间差异不显著外(P>0. 05),其余两代林的各营养元素积累量之间差异显著(P<0. 05)(表2)。 从表2中两代林各养分元素积累量占总养分元素量的百分比(括号内的数)对比中看出,杉木从幼林到成熟期,第一代不同林龄时积累N和P量占总养分量的比例分别比第二代高0.62—2.70,0.03—0.18个百分点,第一代的Ca却比第二代少0.22—1.71个百分点;第一代7年生时的K所占比例比第二代低0.06个百分点,11、16、20、25年生时分别比第二代高0.20—0.39个百分点;虽然11年生时第一代的Mg所占比例比第二代低0.07个百分点,但7、16、20、25年生时第一代比第二代高0.40—1.33个百分点。表明第一代N、P、K各占总养分量比例分别高于第二代,Ca、Mg所占比例少于第二代。 2.3.2 养分在器官分配上的比较 从图3 看出,除7年生时两代杉木林积累的养分在器官分配大小顺序相同外(叶>枝>干>皮>根),7年生以后其顺序发生了变化。11年生林分,各器官积累养分量大小顺序是:第一代叶>枝>干>皮>根,第二代叶>枝>皮>根>干;16年生,第一代干>皮>叶>枝>根,第二代叶>枝>皮>干>根;20年生,第一代干>叶>皮>枝>根,第二代叶>皮>枝 >干>根;25年生,第一代干>皮>叶>枝>根,第二代叶>皮>干>枝>根。 图3 两代杉木林不同生长阶段各器官的养分累积量Fig.3 Nutrient components accumulated in the organs at different growth stages of two generation Chinese fir forests 从幼林到成熟期各林龄时积累在第一代干、皮中养分量比第二代分别多29.79%—79.12%,1.68%—15.21%。积累在第一代的枝、叶养分量除7年生时分别比第二代多1.71%和3.92%外,7年生以后各林龄时则分别比第二代少30.32%—35.56%和7.12%—16.59%。林分16年生以前,积累在根中养分量,第一代比第二代多9.81%—16.36%,16年生以后,第一代比第二代少3.80%—17.10%。 经差异分析得出,除7年生的两代林树皮之间、树枝之间、树叶之间、树根之间,11年生的两代林树皮之间、树叶之间,25年生的两代林树皮之间的养分积累量差异显著(P<0.05)外,其余差异极显著(P<0.01) 通过两代林各林龄时积累养分在各器官中的分配比率对比分析得出:同一林龄时,养分在干、皮中的分配比,第一代比第二代分别高2.99—13.31,0.09—1.79个百分点;枝和叶中分配比,第一代比第二代分别低1.31—8.48,0.62—5.99个百分点;根中的分配比,16年生以前,第一代比第二代分别高0.59—2.23个百分点。16年生以后比第二代少0.70—2.35个百分点。从中看出,第一代积累养分在干和皮的分配比高于第二代,枝和叶中的分配比低于第二代。 2.4.1生产1吨干物质需要的养分 从表3中看出,1—7,8—11,12—16,17—20,21—25年生时,每生产1 t干物质所需养分,第二代比第一代分别多1.28、3.19、4.28、4.09、4.09 kg。就具体营养元素而言,第一代不同生长阶段每生产1 t干物质的需N、P、K、Ca、Mg量,分别比第二代少0.85—1.35,0.05—0.14, 0.51—1.08,0.70—1.50,0.17—0.51 kg。换一种角度分析。假若吸收100 kg养分,以第一和二代不同生长阶段的养分利用效率,在各生长阶段第一代可生产出9.28、7.72、11.60、13.23 t和11.47 t干物质,第二代分别只能生产7.66、6.20、7.75、8.58 t和7.81 t。也就是说不同生长阶段的养分利用效率,第二代比第一代相应低17.46%,19.69%,33.19%,26.03%和31.91%。 表3 两代杉木林不同生长阶段生产一吨干物质所需养分/(kg/ t) 括号内的数为百分数 2.4.2生产1吨杉木器官需要的养分 表4看出,第二代在1—7,8—11,12—16,17—20,21—25 a各生长阶段,生产1 t树干、皮、枝、叶和根的干物质所需养分量,分别比第一代多0.19—0.29,1.38—2.75,2.15—3.36,1.60—3.61,0.88—1.25 kg。各器官养分的利用效率上,第一代比第二代树干的养分利用效率高7.02%—10.22%;皮高11.73%—21.29%;枝高13.12%—21.19%;叶高6.19%—14.03%;根高10.91%—21.43%。 表4 两代杉木林不同生长阶段生产一吨杉木器官需要的养分/(kg/t) 第二代杉木器官养分浓度比同一生长阶段第一代相应的高;进入干材生长阶段后,器官养分浓度随生育阶段的提高而逐渐降低,与马祥庆等[23]的结论相同。本研究第一代杉木林在某一林龄时乔木层养分积累量高于第二代,在另一林龄时稍低于第二代,与马祥庆等[23]和刘爱琴等[22]研究第一代杉木林在任一林龄时乔木层养分积累量都高于第二代结果不一样。本研究表明,杉木连栽并不一定导致第二代乔木层养分积累减少。 不同林龄时养分在器官分配上与马祥庆等[23]研究的有所不同。马祥庆等研究认为虽然不同生长阶段养分在器官分配上表现不一,但同一生长阶段各栽植代数相同。本研究结果除7年生以前两代林养分在器官上的分配大小顺序相同外,8—25年生中的同一生长阶段两代林在器官养分分配大小顺序并非完全一致。马祥庆等的研究地是福建尤溪属中亚热带海洋性季风气候区,会同属中亚热带湿润气候区。同一树种在不同的立地环境条件下,可能造成其林分不同生育阶段生物量在器管分配和器管养分含量的差异,进而导致养分积累在器管分配上的差异。 会同两代林分年均吸收养分量与刘爱琴等[22]研究的也有差别。本研究林分7年生前第一代比第二代多,8—25年生比第二代少。刘爱琴等研究得出,中龄林(第1代11年生,第2代12年生)和成熟林(两代都为31年生)都是第一代多于第二代。而且年均吸收养分量随林龄增加其变化与刘爱琴等的也不一样。刘爱琴等认为随发育阶段的提高,两代林养分的年吸收量都呈增加趋势[22];但会同两代杉木林年均吸收养分量在某个生长阶段均出现下降的趋势。造成两者差别的原因有3点:(1)本研究的两代林是同一立地(第一代采伐后第二年在采伐迹地营造第二代林)生长的林分,刘爱琴研究的不同代数林分,采用“时空转换”法在研究林区内选择林地母岩及地位指数一致的两个不同林分。由于空间异质性,可能使研究的结果与实际并不完全相符。(2)刘爱琴等只测定11年生(或12年生)和31年生两个林龄时,两林龄间相差较大,无法体现出11—31年生其间不同生长阶段吸收养分量变化趋势。(3)刘爱琴等采用“吸收=存留+归还”的方法估算不同生长阶段林分养分吸收量,会同杉木林的不同生长阶段林分养分吸收量是用本文研究方法中的方法估算的。估算方法不同,所得出的结果也就不一样。 第一代杉木的养分利用效率高于第二代,以及养分利用效率随林龄增加的变化上会同的1—20年生,尤溪的1—19年生[23](马祥庆等只研究林龄19年生时)两者趋同。都表明杉木连栽可导致第二代杉木养分利用效率下降。我们曾研究会同两代林的杉木养分利用效率[24](那时只研究到第二代18年生时)。虽然得出的结论与本次相同,但是,此次研究的结果在1—7,8—11、12—16年生时,第一代每生产1 t干物质所需养分量分别比前次研究的第一代多10.85%,28.95%和16.32%,第二代也比前次第二代多4.60%,35.16%和8.63%。造成这种差别是因为前次研究某生长阶段的生产量时,是用某生长阶段测定的生物现存量减去前次生物现存量的差,再加某生长阶段的凋落物量求算的,某生长阶段的养分吸收量是依据“吸收=存留+归还”的计算方法求得。本次研究估算某生长阶段的生产量考虑了枝、叶的实际生长期;估算某生长阶段年均吸收的养分量时不是用“吸收=存留+归还”的方法,而是用考虑了枝、叶的实际生长期后估算的生产量乘以物质的养分浓度,并考虑了这一生长阶段与前一生长阶段生物量中的养分浓度差异,因而这次研究结果更逼近实际。 本研究结果表明,杉木连栽第二代林进入快速生长阶段后年均吸收的养分量比第一代多。这一方面说明为了维持杉木在整个生长过程中的正常生长,第二代林所需的养分比第二代多;而且也表明第二代比第一代从土壤中吸收更多的养分。这对林地养分的消耗大,由此可能导致第二代林地的地力衰退。 任一个生长阶段第二代林分生产力低于第一代(表1)。早就有人报导过植物生长量与养分含量和养分平衡密切相关[3,8]。也就是说当树木体内含有较多的营养元素量,但树木的生长不一定是最好的,只有当这些各营养元素量达到相互平衡时,才有可能获得最好的产量。虽然第二代杉木林体内养分含量比第一代的高,但通过对杉木体内各养分元素量搭配的比例分析(表2中括号内的数据),第一代1—7,8—11,12—16,17—20,21—25年生时杉木体内的N、P、K、Ca、Mg占总养分的比例与第二代不相同。如果认为第一代杉木体内各元素量的比例相对平衡有利杉木生长的话,那么第二代的各元素量比例的不协调可能是导致第二代养分利用效率降低,以及生产力下降的一个主要原因。究竟是什么原因引起两代杉木体内各元素量搭配的比例不相同,养分利用效率下降,有待进一步深入研究。 从本研究结果看出连栽的第二代林的养分吸收量比第一代多,而养分利用效率低于第一代。这些都有可能导致第二代林的生产力降低和地力衰退。若不解决好这些问题人工林生产经营将很难持续。因此必须在营林措施上下功夫。如采用混交林栽植模式可以改变纯林的单一结构导致人工林生态系统养分循环失调[26],采用施肥、抚育和不同的采伐强度等经营管理措施可提高人工林土壤肥力、缓解产量下降[27]。人工林地力衰退可以通过改变人工林经营管理方式,保证人工林生长的一个良性循环的生态环境,人工林连栽是可行的[28-29]。 本文研究是在第二代与第一代连栽的林地进行的。避免了“时空转换”法的缺陷,保证了空间尺度的一致性和时间尺度的连续性。在不同生长阶段吸收的养分量估算上采用与以往同类研究中完全不相同的方法。这种研究方法真实地反映了杉木生长和对养分吸收利用的生理规律。 (1)同一生育阶段第二代杉木器管的各营养元素浓度都比第一代相应的高。其中,N浓度高2.85—3.48 g/kg,P浓度高0.16—0.25 g/kg,K浓度高1.86—2.72 g/kg,Ca浓度高2.10—2.50 g/kg,Mg浓度高0.77—1.31 g/kg。 (2)第一代杉木林在某一林龄时乔木层养分积累量高于第二代,在另一林龄时也可能低于第二代,杉木连栽不一定导致第二代林乔木层养分积累减少。养分在各器官中的分配比两代林中不相同。第一代养分在树干的分配比高于第二代,枝和叶中的分配比低于第二代。 (3)年均吸收养分量,7年生以前,第一代比第二代多,7年生以后,第二代比第一代多。表明杉木连栽第二代林会从土壤吸收更多的养分,不利于土壤肥力水平的维持 (4)任一个生育阶段,生产1吨杉木干物质所需养分量,第一代少于第二代,而且生产1吨干、皮、枝、叶和根干物质所需的养分量,也是第一代少于第二代。第二代的养分利用效率比第一代低,这应是杉木连栽第二代杉木林生产力下降的原因之一。1.6 养分利用效率
1.7 数据分析处理
2 结果与分析
2.1 养分浓度的比较

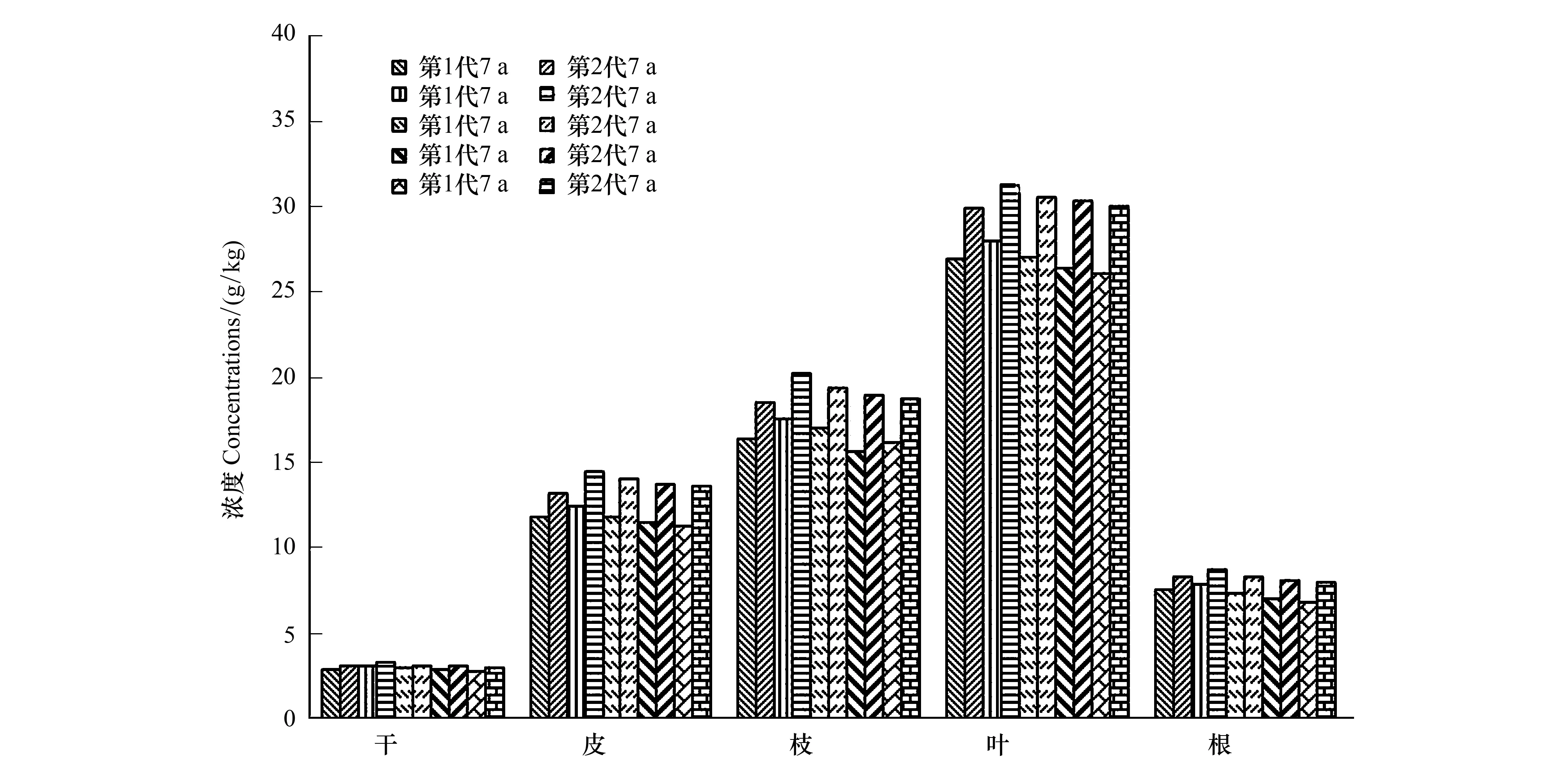
2.2 两代杉木林年均吸收养分的比较
2.3 养分积累与分配的比较

2.4 养分利用效率比较
3 讨论
4 结论
