Changes in fi sh diversity and community structure in the central and southern Yellow Sea from 2003 to 2015*
2018-07-11CHENYunlong陈云龙SHANXiujuan单秀娟JINXianshi金显仕JOHANNESSENArneYANGTao杨涛DAIFangqun戴芳群
CHEN Yunlong (陈云龙) SHAN Xiujuan (单秀娟) JIN Xianshi (金显仕) JOHANNESSEN Arne YANG Tao (杨涛) DAI Fangqun (戴芳群)
1Institute of Oceanology,Chinese Academy of Sciences,Qingdao 266071,China
2Function Laboratory for Marine Fisheries Science and Food Production Processes,Qingdao National Laboratory for Marine Science and Technology,Qingdao 266237,China
3Key Laboratory of Sustainable Development of Marine Fisheries,Ministry of Agriculture,Shandong Provincial Key Laboratory of Fishery Resources and Ecological Environment,Yellow Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Qingdao 266071,China
4University of Chinese Academy of Sciences,Beijing 100049,China
5Department of Biology,University of Bergen,PO Box7803,5006 Bergen,Norway
AbstractThe central and southern Yellow Sea is an important overwintering ground for many fi sh species in the Bohai Sea and Yellow Sea. For better understanding the status of the fi sh community after years of heavy exploitation, variations in fi sh community structure and diversity were analyzed using data from bottom trawls during 2003-2015. Five fi sh assemblage indices all showed fluctuations without clear trends from 2003 to 2015, yet there were strong positive and significant correlations (P <0.05) among them.The top-fi ve dominant species accounted for a high weight percentage (49.7%-82.1%) in the annual fi sh catch. Multivariate analysis showed that two year groups could be pooled for the fi sh community: Group I consisted of the years 2006, 2007, 2008 and 2015, while Group II consisted of the years 2003, 2004, 2005,2009, 2010 and 2014; the groups aggregated with 63.71% similarity, indicating a high level of similarity among all years. The multivariate dispersion values were 1.455 and 0.818 for Groups I and II, respectively,indicating greater variances in fi sh assemblage structure in Group I than that in Group II. Similarity of percentage analysis demonstrated that the average similarities for Group I and Group II were 71.58% and 67.51%, respectively. Size-spectra analysis revealed no consistent trend in the intercept and slope (P >0.05);there were also no significant differences between the slope of the size-spectra and fi shing effort. The catch per unit effort and mean individual weight analyses of the whole fi sh assemblage both showed a significantly decreasing trend over time. Overall, the results showed that the fi sh community structure in the central and southern Yellow Sea was relatively stable from 2003 to 2015 and the study could be used as a reference for supporting ecosystem-based fishery management.
Keyword:dominant species; fi sh community structure; multivariate analysis; trawl data
1 INTRODUCTION
The Yellow Sea is a marginal sea located in the western Pacific Ocean between China and the Korean Peninsula with an average depth of 44 m. It is shallow along the coast and becomes relatively deep in the Yellow Sea trough area (Wei et al., 2010; Lie et al.,2015). In winter, the Yellow Sea Warm Current is the most remarkable hydrological event in the Yellow Sea. It carries seawater of high temperature and salinity from the northward Kuroshio branch current and the Taiwan Warm Current in the northern part of the East China Sea and intrudes into the central Yellow Sea (Lin et al., 2011). Dominated by winter monsoons, several southward coastal water currents which are usually cold, full of nutrients and have low salinity move to the central Yellow Sea (Chen, 2009).Together with discharge from the Changjiang River,these water masses interact in winter and drive the biogeochemical cycles in this region. Especially, they determine the hydrological environment, matter transportation and sedimentation, which have great impacts on the distribution of plankton, fishes,crustaceans and cephalopods (Qin et al., 1989; Jin et al., 2003; Liu et al., 2015; Mei et al., 2016).
As an important ecological unit of spatial scale,fi sh community reflects the characteristics of the fishery ecosystem from the aspects of material recycling and energy transformation. The maintenance of biodiversity and the stabilization of fi sh community are critical to the balance of aquatic ecosystems.Many external factors have been proven to inf l uence fi sh communities such as overfishing, reclamation,pollution and climate change, which may lead to lower fi shing rates, the miniaturization and early maturation of fi sh stocks (Dethlefsen and Tiews,1985; McLusky et al., 1992; Jennings et al., 2001;Genner et al., 2010; Stefansdottir et al., 2010).
Fisheries resources research in the Yellow Sea has been widely conducted and has mainly concentrated on species composition, population dynamics and fi sh community structure (Jin and Tang, 1996; Jin, 2003;Xu and Jin, 2005; Li et al., 2011; Shan et al., 2012;Wang et al., 2013; Huang et al., 2014). Notably, Xu and Jin (2005) studied variations in the fi sh assemblage structure over the period 1985-2002. However, since then, the fishery environment has changed with variations in the ecological environment and everincreasing fi shing efforts (Keesing et al., 2011; Yuan et al., 2012). The exploitation of Japanese anchovyEngraulisjaponicusTemminck & Schlegel 1846, a key species in the Yellow Sea ecosystem, epitomizes the fate of many economically important species.After heavy exploitation of Japanese anchovy since the 1990s, its biomass fluctuated at low levels, ranging from 0.11 to 0.42 million tons during 2001-2005(Zhao, 2006). Considering this background, what subsequent changes are to be expected for the fi sh community in the central and southern Yellow Sea?
Fig.1 Schematic of the sampling stations in the central and southern Yellow Sea
In this study, fi sh assemblage indices were calculated to assess variations in the fi sh community diversity in the central and southern Yellow Sea for the period 2003-2015. Dominant species composition was explored to determine the characteristics of the fi sh community during this period. Multivariate analysis and analyses of mean individual weight(MIW), size-spectra, and catch per unit effort (CPUE)were used to advance information on the whole fi sh community during the sample years. Reasons for variations in the fi sh community in the region are also discussed. This research benefi ts assessments of the fi sh community in the Yellow Sea and provides data that support ecosystem-based fishery management in the future.
2 MATERIAL AND METHOD
2.1 Survey area and data collection
Sampling stations were designed on a grid with regular intervals of 0.5°N×0.5°E (Fig.1). Information on survey times and the number of bottom trawls from 2003 to 2015 (excepting 2011-2013) are listed in Table 1. Although the number of survey stations varied between years, relatively high sampling coverage was performed annually throughout the entire geographic range. Furthermore, a bootstrap method (Felsenstein, 1985) was used to balance the uneven effect of trawl numbers.
Data were collected by the Yellow Sea Fisheries Research Institute in winter during the period 2003-2015, using the R/VBeidou. Parameters of the fi shing gear were as follows: a net of circumference 836 mesh×12 cm, and a 10-cm mesh-size cod-end with a 2.4-cm mesh-size liner thereafter. The duration of each trawl haul varied between 0.5 h and 1 h at an average hauling speed of 3 knots. All data were standardized to 1 h trawl duration for further analyses.The catch at each station was identified to species level, or to the lowest possible taxon level, and the abundance (in ind.) and weight (in g) of each species were recorded. Fishing effort (in hp) was calculated by the sum of total horsepower of mobile fi shing vessels, including those from Shandong Province,Hebei Province, Liaoning Province and Tianjin City using China fi sheries statistics yearbook.
2.2 Data analysis
2.2.1Fish assemblage indices
Fish assemblage indices in this study consisted of three types: indices of diversity, richness and evenness. The indices were defi ned using relative biomass (in kg/h) to avoid the dissimilarity caused by different individual sizes (Qiu, 1988; Jin and Tang,1996). For diversity indices, the Shannon-Wiener index (H') and Hill’s family of diversity numbers (N1andN2) were used (Wilhm, 1968; Hill, 1973).N1represents the number of common species, andN2denotes the number of dominant species in the fi sh community. The Margalefindex (d) was chosen to represent the richness of the fi sh community due to its independence from sample size, and Pielou’s evenness index (J') was calculated to determine the evenness of a community (Margalef, 1958; Pielou, 1975; Jin and Tang, 1996). Based on the catch data in weight obtained by bottom trawls from 2003 to 2015, the diversity, richness and evenness of the fi sh assemblages were calculated to a 95% confi dence interval. The calculation process of each index was repeated 1 000 times for each year using a bootstrap method.
2.2.2Multivariate analysis
A fourth-root transformation was fi rst applied to the biomass data, following the method of Xu and Jin(2005), so as to reduce the effect of dominant species on the ordination, using the Bray-Curtis similarity measure. Hierarchical clustering and non-metric multidimensional scaling (NMDS) were used to analyze the composition characters of the fi sh assemblages between the survey years. A stress value was calculated to evaluate the difference between the distance in the reduced dimension and the complete multidimensional space: a stress score below 0.1 is considered good, while a stress score above 0.2 is considered unacceptable (Clarke, 1993). The multivariate dispersion (MVDISP) index was used to reflect the amount of variation in the fi sh assemblage structure under abiotic and biotic disturbances indifferent year groups. Similarity percentage(SIMPER) analysis was performed to examine both the contribution of each species to the average Bray-Curtis dissimilarity between different year groups and the similarity within each year group (Clarke, 1993;Clarke and Warwick, 2001; Xu and Jin, 2005).
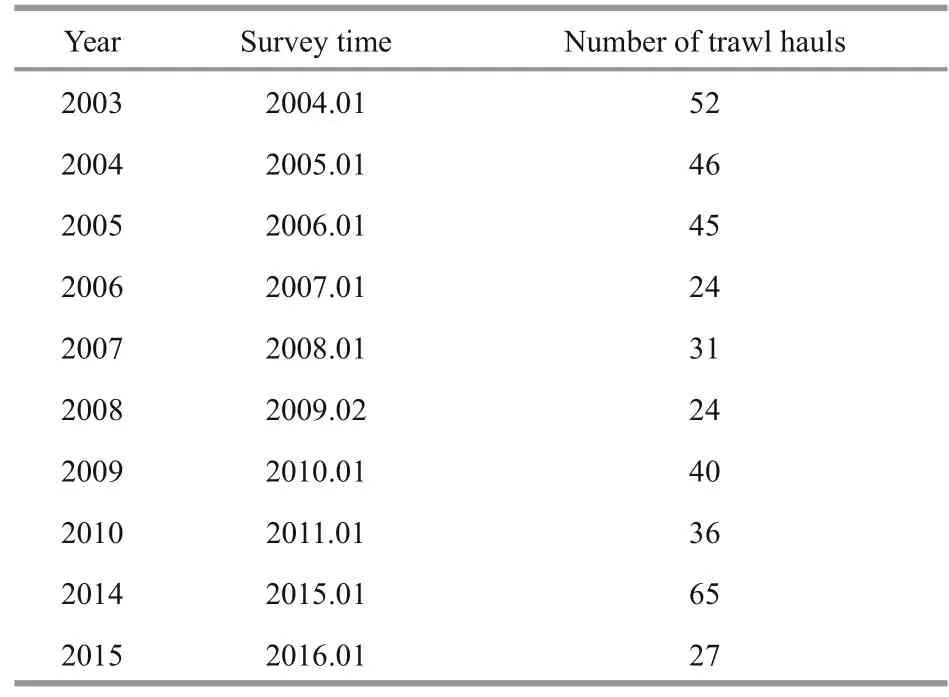
Table 1 Summary of the trawl surveys in the central and southern Yellow Sea
2.2.3Size-spectra, MIW and CPUE analyses
In size-spectra analysis, the average weight (in g)of each species was fi rst calculated by its corresponding weight and the number ofindividuals captured at each sampling station in each year. Next, this average weight was converted into average length (in cm),based on the hypothesis that length=(100× weight)1/3,and then the number ofindividuals captured per hour was assigned to a corresponding length group (Xu and Jin, 2005). The size-spectra were plotted using the summed length of all species as an independent variable, with intervals of 5 cm, and the natural logarithm of summed individual numbers of all species as the dependent variable. Linear regressions were performed for fi sh lengths ranging from 5 to 65 to estimate the slopes and intercepts of the sizespectra (Gislason and Rice, 1998; Bianchi et al., 2000;Xu and Jin, 2005). Species occurrence of more than three times and seven times were included in the MIW and CPUE analyses as low and high thresholds,respectively. The MIW of each species was calculated by dividing the corresponding total weight by the total individual numbers annually, while the CPUE of each species was calculated by dividing the total individual number by sampling stations per year. The average weight and CPUE for each species and their relationship with years was explored by Spearman’s rank correlation. TheG-test was used to test forsignifi cance of size changes over time, andG-statistics were obtained by the signs frequency of Spearman’s rank correlation coeffcients (Zar, 1984; McDonald,2014).
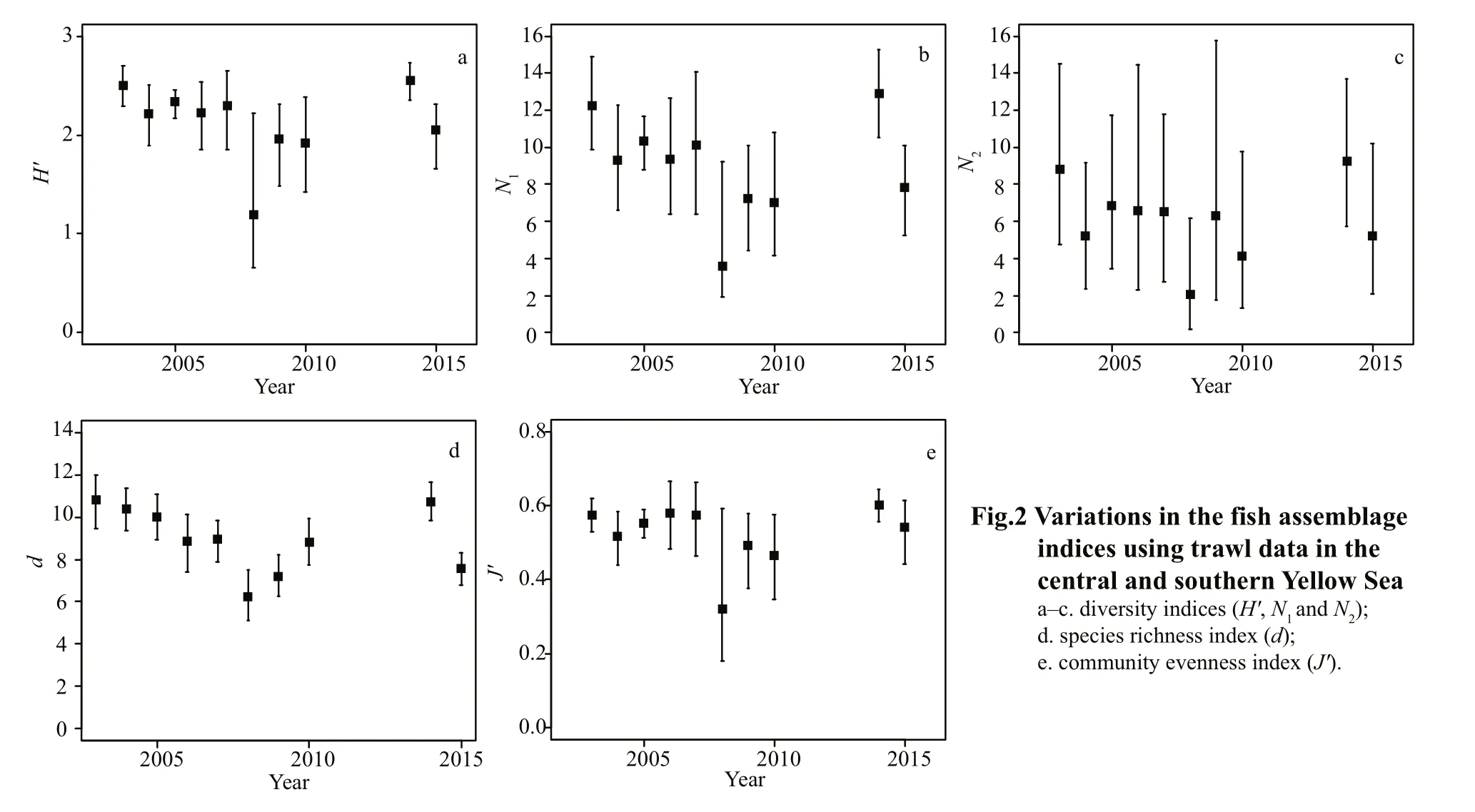
Fig.2 Variations in the fish assemblage indices using trawl data in the central and southern Yellow Seaa-c. diversity indices (H′,N1 andN2);d. species richness index (d);e. community evenness index (J′).
Fish assemblage indices and multivariate analysis were performed with PRIMER v.5.2.8 software(Clarke and Gorley, 2001; Clarke and Warwick,2001); the other analyses namely size-spectra, MIW and CPUE, were performed using R v.3.3.0 (www.rproject.org; Ihaka and Gentleman, 1996).
3 RESULT
3.1 Variations in the fi sh assemblage indices
All five fi sh assemblage indices showed fluctuations but without any clear trends for 2003-2015 (Fig.2)(Spearman’s rho=-0.33 forH', Spearman’s rho=-0.33 forN1, Spearman’s rho=-0.22 forN2, Spearman’s rho=-0.52 ford, Spearman’s rho=-0.18 forJ'; allP>0.05). However, the Shannon-Wiener index (H'),N1, and the Margalefindex (d) revealed a significant decreasing trend between 2003 and 2010 (Spearman’s rho=-0.76 forH', Spearman’s rho=-0.76 forN1,Spearman’s rho=-0.88 ford; allP<0.05), with the lowest values resulting from the year 2008 (Fig.2).The results ofN2and the Pielou evenness index (J')showed a decreasing trend during the same period,but this was not significant (Spearman’s rho=-0.67,P=0.07 forN2; Spearman’s rho=-0.64,P=0.09 forJ').
There were strong positive and significant correlations (P<0.05) among the fi sh assemblage indices (Fig.3), and all five indices displayed the same trend. The highest correlation was found between diversity indicesH'andN1( Pearson’s rho=0.97),while the lowest correlation was found between the richness index and the evenness index (Pearson’s rho=0.72).
3.2 Changes in dominant species between years
The number of dominant species for a given year ranged from 2 to 9 during 2003-2015. In 2008, the Japanese anchovy and the anglerfishLophiuslitulon(Jordan 1902) accounted for 75.4% and 6.7% of the total fi sh catch respectively. Japanese anchovy and the anglerfish were found to be the dominant species in all survey years (10 yrs) followed by Tanaka’s snailfishLiparistanakae(Gilbert & Burke 1912)(dominant in 9 yrs), silver pomfretPampusargenteus(Euphrasen 1788) (8 yrs) and small yellow croakerLarimichthyspolyactis(Bleeker 1877) (5 yrs). These top-fi ve species ranged from 49.7% to 82.1% in weight percentage of the total fi sh catch from 2003 to 2015. Four lesser dominant species (occurrence fewer than 4 yrs) were smallhead hairtailEupleurogrammus muticus(Gray 1831) (3 yrs), Pacific codGadus macrocephalusTilesius 1810 (2 yrs), scaly hairfi n anchovySetipinnataty(Valenciennes 1848) (2 yrs)and Osbeck’s grenadier anchovyCoiliamystus(Linnaeus 1758) (2 yrs). Total weight of all dominant species accounted for a large proportion of the total fi sh catch, ranging between 76.1% and 89.0 %.
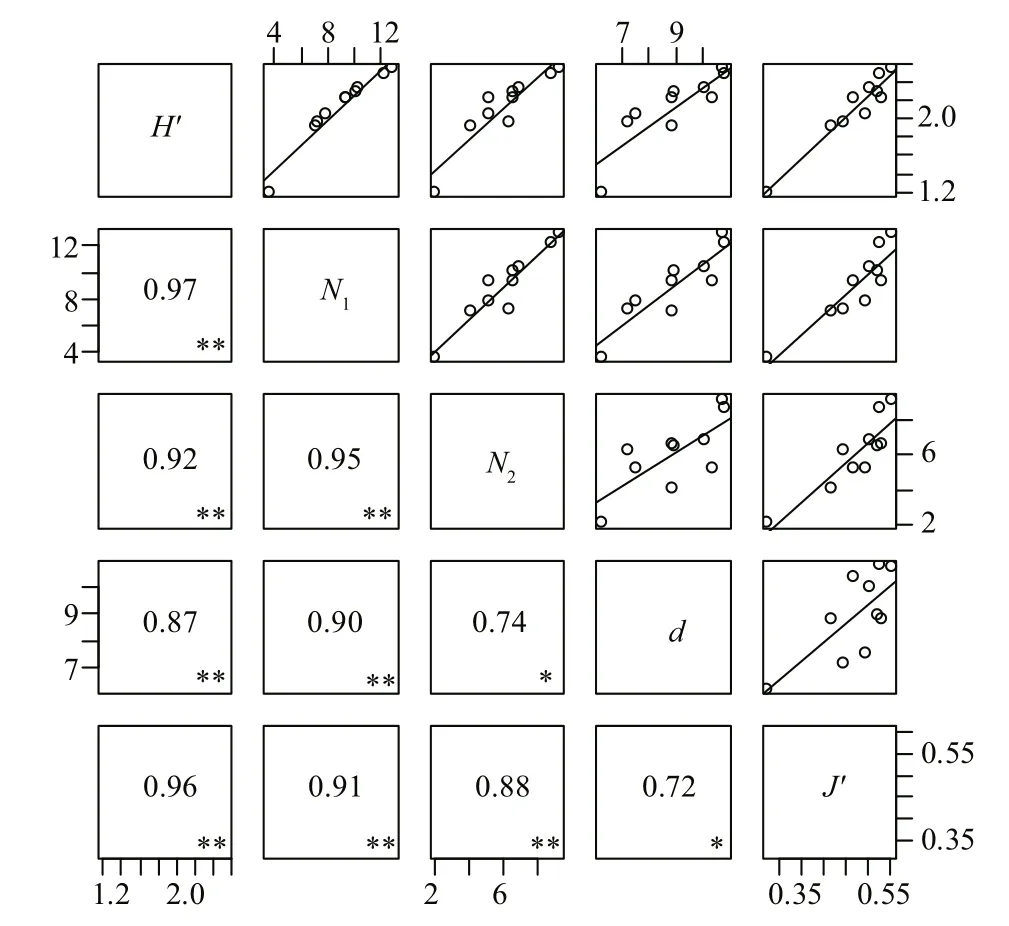
Fig.3 Pearson correlation coeffcients between the fi sh assemblage indices (* P< 0.05, ** P< 0.01)
3.3 Multivariate analysis
Cluster analysis of fi sh assemblages in the central and southern Yellow Sea was conducted using Bray-Curtis similarity between the years (Fig.4). Two groups (Groups I and II) were identified and aggregated with 66.16% and 69.36% similarity,respectively. However, the two groups aggregated together with 63.71% similarity, which means there was a high level of similarity among all years.
The stress score of the NMDS analysis was 0.06(<0.1), indicating that the composition of species from different years could be accurately distinguished.In Fig.5, Group I is localized in the left-upper region of the fi gure and Group II in the central and right region. The NMDS analysis was in accordance with the results of the cluster analysis.
The MVDISP values were 1.455 and 0.818 for Group I and Group II, respectively, which indicated greater variances in the fi sh assemblage structure in Group I than that in Group II during the sample period. This was in accordance with the results of the NMDS analysis in which the distances of the survey years within Group I were slightly farther apart from each other as compared to the distances within Group II, which were much closer.

Fig.4 Cluster analysis of the fi sh assemblages from trawldata in the central and southern Yellow Sea▲ Group I; ■ Group II.
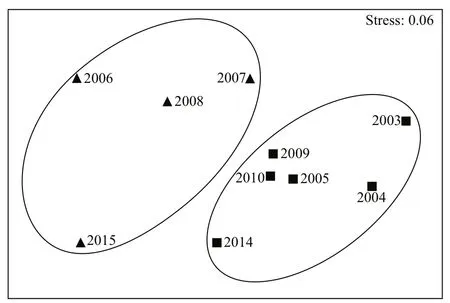
Fig.5 Non-metric multi-dimensional scaling (NMDS)analysis in the central and southern Yellow SeaStress value=0.06, ▲ Group I, ■ Group II.
The SIMPER analysis demonstrated that the average similarities for Group I and Group II were 71.58% and 67.51%, respectively. Twelve species typified in both groups including the anglerfish, Japanese anchovy,Tanaka’s snailfish, silver pomfret, Pacific cod, small yellow croaker, smallhead hairtail, scaly hairfi n anchovy, plaiceCleisthenesherzensteiniSchmidt 1904, bartail flatheadPlatycephalusindicus(Linnaeus 1758), spotted velvetfishErisphexpottii(Steindachner 1896) and whitespotted congerCongermyriaster(Brevoort 1856), they accounted for 44.63% and 53.21% of similarity in Group I and Group II,respectively (Table 3). Additionally, Dotted gizzard shadKonosiruspunctatus(Temminck & Schlegel 1846), chub mackerelScomberjaponicusHouttuyn 1782, Mi-iuy croakerMiichthysmiiuy(Basilewsky 1855), bluefi n gurnardChelidonichthyskumu(Cuvier 1829)and Korean rockfishSebastes schlegeliiHilgendorf 1880 were typifying species in Group I while big head croakerCollichthyslucidus(Richardson 1844),belanger’s croakerJohniusbelangerii(Cuvier 1830),Japanese seabassLateolabraxjaponicas(Cuvier 1828)and other three species were typifying species in Group II.
The dissimilarity was 36.29% between Group I and Group II, which was in accordance with the result of cluster analysis above (Fig.4). The top fi fteen species contributing to dissimilarity between groups were showed in Table 4. Percentage contributions of Japanese anchovy, Tanaka’s snailfish, silver pomfret and Pacific cod were higher than 2% while those of others were less than 2%.
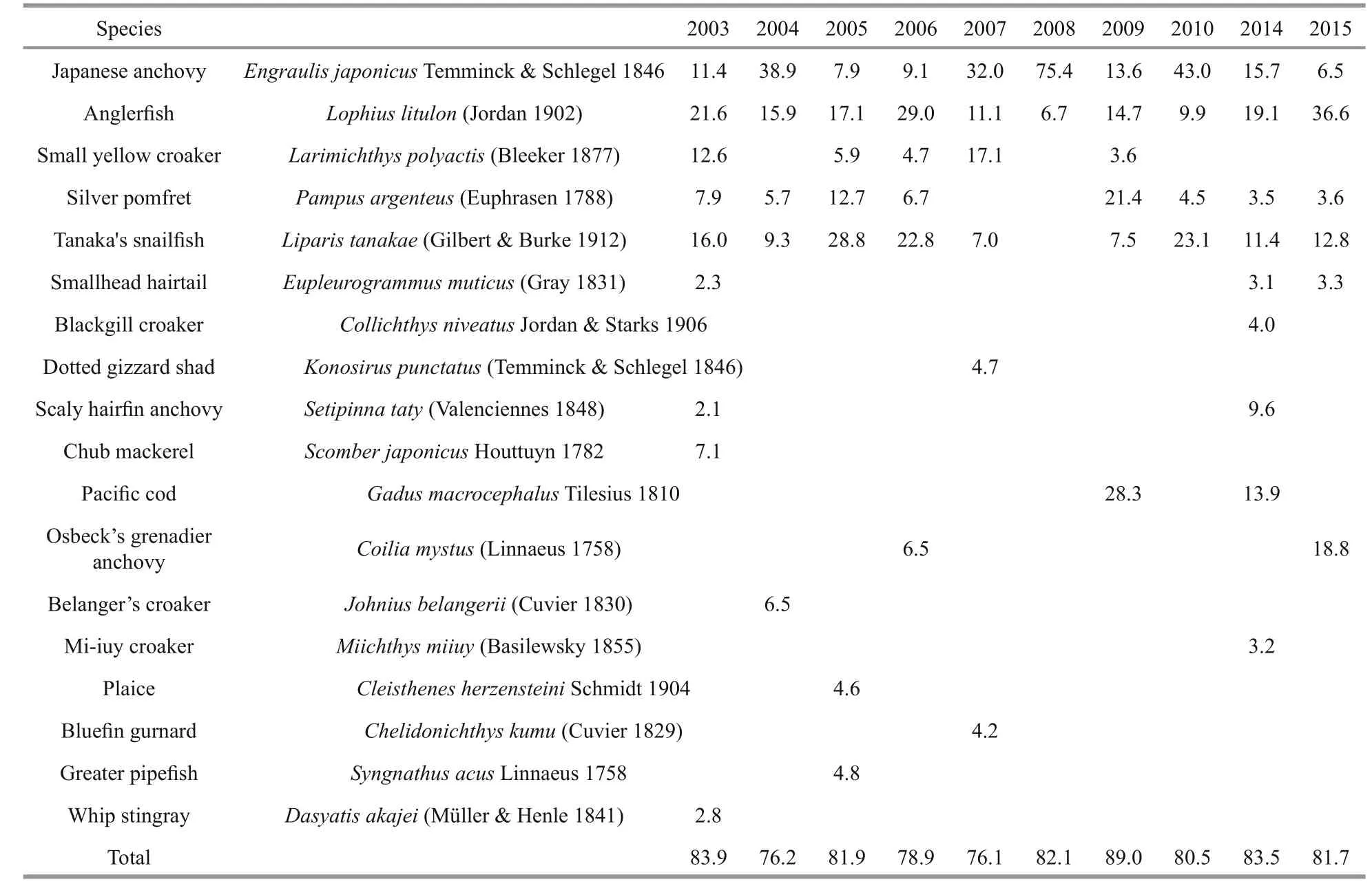
Table 2 Composition of the dominant species and their weight percentages in the total fi sh catch during the survey years in the central and southern Yellow Sea
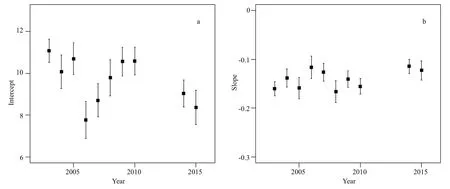
Fig.6 Variations in intercepts and slopes of the whole fi sh assemblage size-spectra based on trawl data in the central and southern Yellow Sea
3.4 Size-spectra, MIW and CPUE analyses
Referring to size-spectra analysis, no consistent trend was found in the intercept (Spearman’s rho=-0.43,P>0.05) and slope (Spearman’s rho=0.42,P>0.05) over time (Fig.6). Intercepts between Group I and Group II were significantly different (F1,8= 11.48,P<0.01), while the slopes were not (F1,8=0.85,P>0.05). There were also no significant differences between the slope of size-spectra and fi shing effort(Spearman’s rho=0.62,P>0.05,n=10).
Species occurring more than seven times during the survey years (high threshold level) were selected for analyses of MIW and CPUE. The MIW of the selected species showed a decreasing trend over time since the number of species with a negative Spearman’s rank correlation coeffcient was twice that of species with a positive value (Table 5). TheG-test indicated that the present distribution of MIW was significantly different from the expected 50:50 null hypothesis (G=4.757,P<0.05).
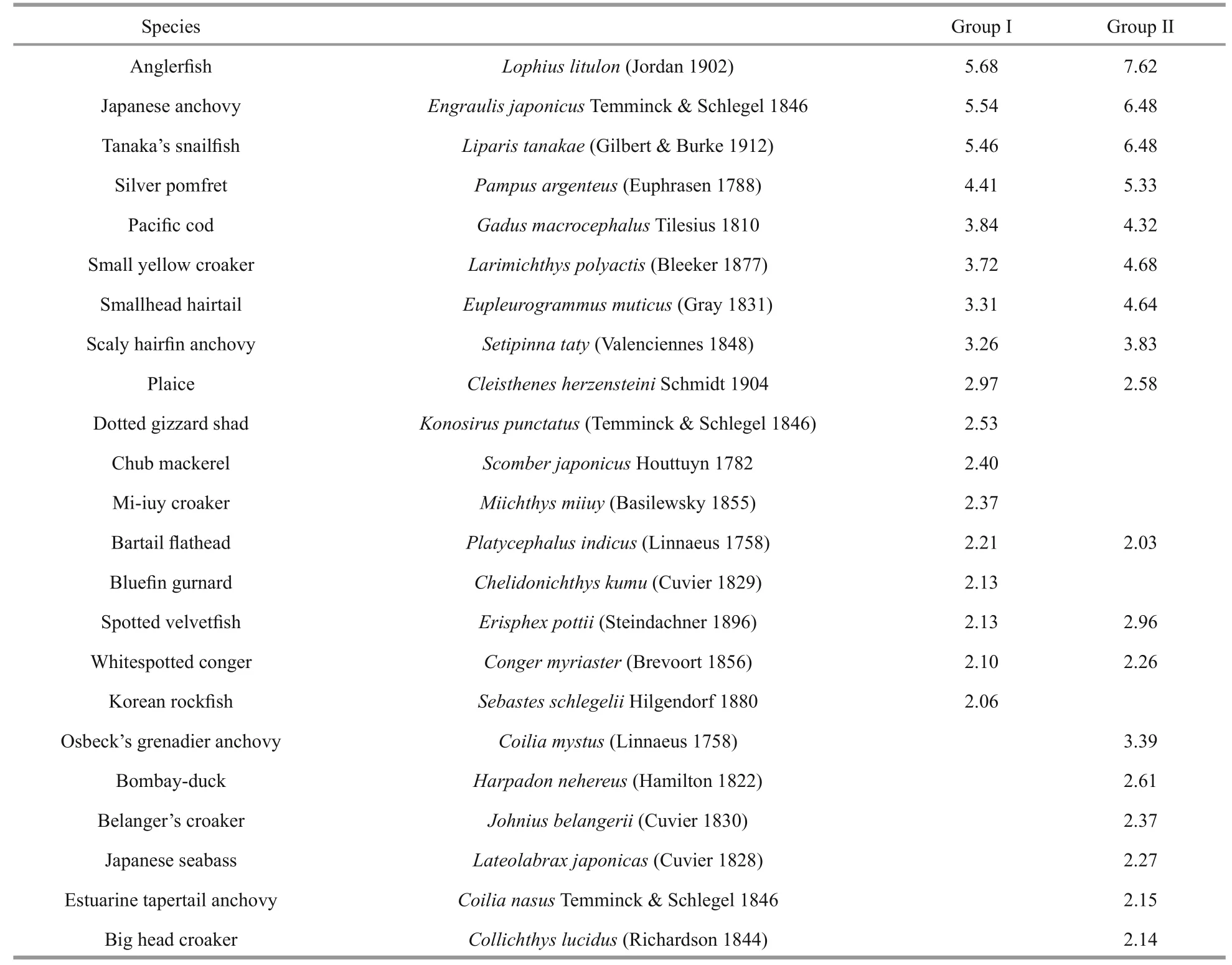
Table 3 Species contributing to similarity (>2%) within groups, as calculated by SIMPER analysis

Table 4 Species most contributing to dissimilarity between groups, as calculated by SIMPER analysis
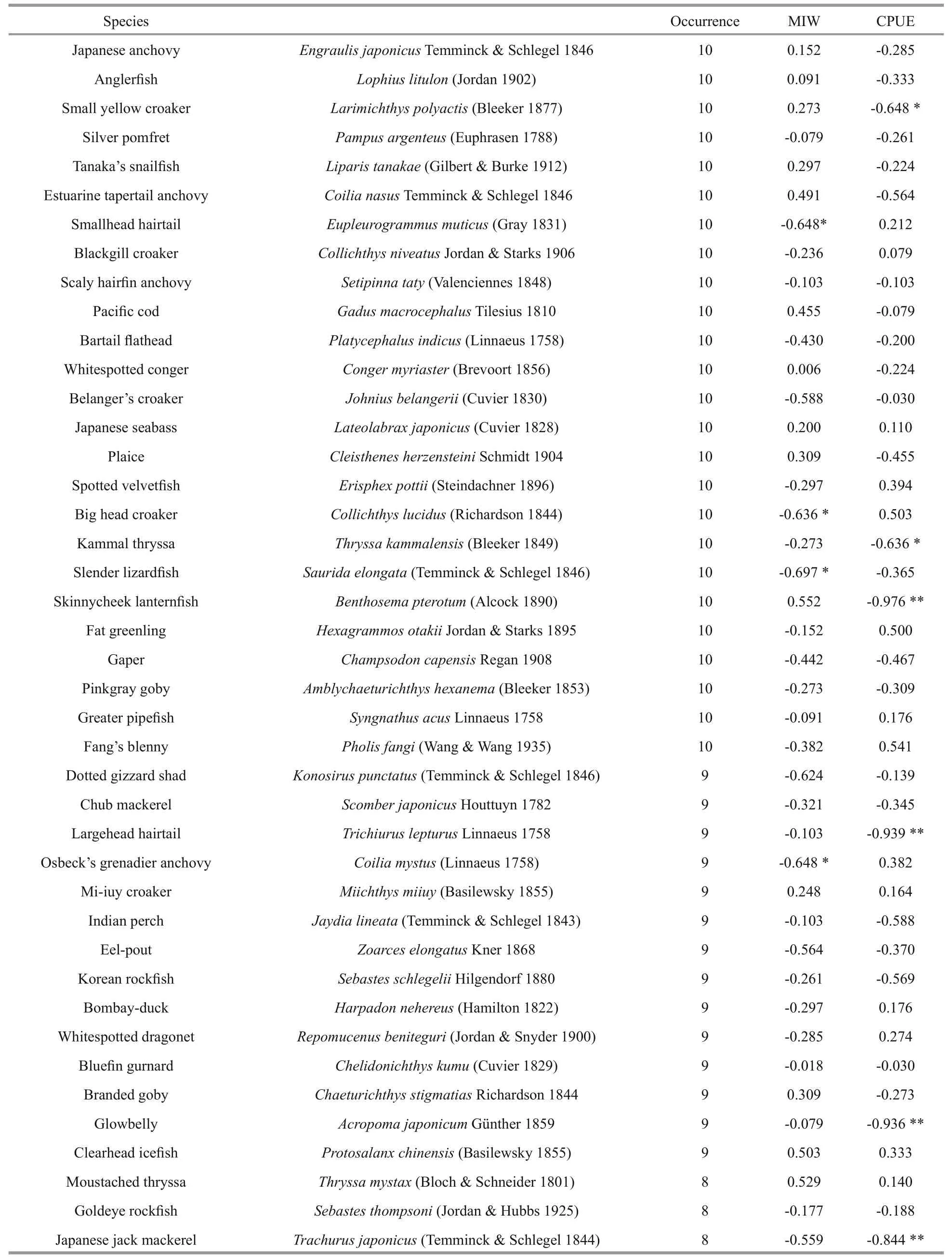
Table 5 Occurrence (years in the trawl data) and Spearman’s rank correlation coeffcients of mean individual weight (MIW)and catch per unit effort (CPUE) with year in the central and southern Yellow Sea
The results of the CPUE analysis were consistent with the MIW analysis and likewise showed a significant decreasing trend over time (G=4.757,P<0.05). The individual correlation coeffcients of some traditional economically important species,such as small yellow croaker and largehead hairtail were significantly high, reaching -0.648 and -0.939,respectively. This phenomenon was likewise observed for some small-sized species, for example, Kammal thryssaThryssakammalensis(Bleeker 1849),skinnycheek lanternfishBenthosemapterotum(Alcock 1890), and glowbellyAcropomajaponicumGünther 1859.
To make the results more persuasive, species occurring more than three times (low threshold level)were also tested. The MIW (G=12.199,P<0.01) and CPUE (G=7.587,P<0.01) again revealed a significant decreasing trend over time.
4 DISCUSSION
As the Yellow Sea is a typical marginal sea, the fishery resources in this region have an independent character. For example, most fishes inhabit the area throughout their life, and thus form relatively independent groups. Their biology and ecology are similarly inf l uenced by the seawater temperature: fi sh overwintering in the central and southern Yellow Sea migrate to coastal waters in spring to spawn with the rise in water temperature, feed in summer and autumn,and then they move back again to the central and southern Yellow Sea for overwintering as the temperatures begin to decrease in winter (Deng and Zhao, 1991; Jin and Tang, 1996). In this study, data for winter was chosen to cover the effect that population migration may or may not have on the fi sh community.
4.1 Fish assemblage indices
Five fi sh assemblage indices all showed similar trends, with significant (P<0.05) correlations between each other, but no clear trends for the entire period 2003-2015. However, a decreasing trend was observed in all five indices for the shorter period 2003-2010, and this was significant (P<0.05) in the Shannon-Wiener index (H'),N1, and Margalefindex(d), but not inN2and the Pielou evenness index (J').This result is consistent with many other research fi ndings, showing that fi sh assemblage evenness may not necessarily be sensitive to variations in environmental conditions (Zhu et al., 1994; Liu et al.,2007; Kamrani et al., 2016). All indices reached a minimum with the year 2008, which might be attributable to a huge biomass of Japanese anchovy which accounted for up to 75.4% of the total fi sh catch that year, while in the other years it contributed less than 50%. Fish assemblage indices should be used carefully in assessing the ecological status classifi cation of fishery resources, as they can be inf l uenced by many salient elements, such as sampling methods, sampling size, and bottom sediment type(Salas et al., 2006; Taherizadeh and Sharifi nia, 2015;Kamrani et al., 2016).
4.2 Fish community structure
Both anthropogenic activities and environmental variables can inf l uence variations in coastal fi sh communities (Olsson et al., 2012; Bergström et al.,2016). For instance, temperature, dissolved oxygen,pH and salinity were the four main variables inf l uencing the fi sh community structure in Iranian coastal waters (Kamrani et al., 2016). However, one challenge in the present study was a lack of accompanying environmental data, which future studies should try to incorporate.
Fisheries resources in the Yellow Sea have seriously declined during the last 50 years, with researchers reporting alternations to dominant species composition, miniaturization of size classes, and trophic level decreases (Jin and Tang, 1996; Jin, 2003;Xu and Jin, 2005). Taxonomic indices, such as average taxonomic distinctness Δ+and variation in taxonomic distinctness Λ+, were both found to be lower in 2000 as compared to 1985 (Xu et al., 2005).The trophic level estimation of high-trophic-level communities in the Yellow Sea dropped from 3.7 in 1985-1986 to 3.4 in 2000-2001, decreasing by 0.14 on average per decade, and the estimation for the northern Yellow Sea was 0.24 per decade from 1985 to 2010 (Zhang and Tang, 2004; Zhang et al., 2007,2011a). Several commercially important species, such as small yellow croaker, largehead hairtail and plaice,were once dominant in the 1950s; Pacific herringClupeapallasiiValenciennes 1847 became abundant in the early 1970s; the small-sized pelagic species of Japanese anchovy, scaly hairfi n anchovy, and Japanese sardinellaSardinellazunasi(Bleeker 1854) fl ourished in the 1980s; Japanese anchovy was found to be the most abundant species in the Yellow Sea in 1986, but thereafter a targeted fishery rapidly developed (Jin and Tang, 1996; Zhao et al., 2003). The high level of exploitation exceeded the resilience of the Japanese anchovy population, leading to a dramatic drop in the stock, from 2.16 million tons in 1987 to 0.18 million tons in 2002 (Zhao et al., 2003).
In the present study, we found that the dominant species composition had become more-or-less equally distributed compared to the overwhelming dominance of Japanese anchovy during 1985-2002 (Xu and Jin,2005). Japanese anchovy still appears to be a key species in the central and southern Yellow Sea, its weight percentage was not as high as prior to 2000,although it showed a surge in 2008. Meanwhile, the weight percentages of the anglerfish and Tanaka’s snailfish rose, and together with Japanese anchovy,these species formed a combination that commonly dominated the whole fi sh community, with their individual percentages ranging from 35.7% to 82.1%(2003-2015). The weight percentages of silver pomfret fluctuated slightly between the sample years;small yellow croaker was a dominant species before 2010; and, Pacific cod has recovered to some degree since 2009. Other species like smallhead hairtail,scaly hairfi n anchovy, Osbeck’s grenadier anchovy and chub mackerel performed occasionally as dominant species.
Many factors likely contributed to the rise in weight percentages of the anglerfish and Tanaka’s snailfish. First, both species have wide feeding habits.The prey species of the anglerfish are numerous and vary based on habitats; this lessens the possibility for drastic changes of the anglerfish resource from the fluctuations of single prey species (Zhang et al.,2010); Tanaka’s snailfish is omnivorous, has a high feeding intensity, and probably has the highest rate of food consumption among the Yellow Sea fishes (Xue et al., 2007). Secondly, top carnivorous species such as Spanish mackerelScomberomorusniphonius(Cuvier 1832), whitespotted conger, and largehead hairtail had been replaced by the anglerfish and Tanaka’s snailfish whose trophic level reaching 4.4 and 4.2, respectively, in an analysis of the Yellow Sea’s food web (Zhang and Tang, 2004). Fluctuations in populations of top predators can greatly inf l uence the abundance of other species through the top-down effect (Scheffer et al., 2005). Thirdly, the anglerfish and Tanaka’s snailfish both benefi t as a result of their particular reproductive strategies. Anglerfish produce egg veils, which aids fl otation, nutrition support and fertilization of the eggs (Armstrong, 1992; Zhang et al., 2011b). No significant differences (P>0.05) were found in the fecundity of Tanaka’s snailfish in the 2010s compared with those in the 1980s, indicating that the species has a high degree of adaptation to the increasing fi shing effort (Shan et al., 2014).Considering the declines in traditional economically important species and Japanese anchovy in particular,the biology of these fast-growing species makes them highly competitive and well-adapted to occupy vacant ecological niches.
4.3 Multivariate analysis
Multivariate analysis demonstrated that the survey years could be divided into two groups. However, it is diffcult to explain why these Specific years grouped together because fi sh assemblages are inf l uenced by both biotic and abiotic elements, which are complex(Taherizadeh and Sharifi nia, 2015). A challenge to the research was a lack of environmental data collected along with the trawl data. Although Group I and Group II could be well separated by the NMDS and cluster analyses, the survey years all displayed a high level of similarity, reaching 63.71%.Interestingly, the partitioning of groups in the present study was not aligned over time, which was different from what was observed by Xu and Jin (2005). We propose that this may reflect the change of marine ecosystem after the large-scale exploitation of the Japanese anchovy.
4.4 Size-spectra, MIW and CPUE analyses
No significant differences between the slope of size-spectra and fi shing effort (Spearman’s rho=0.62,P>0.05,n=10) were found, although fi shing effort steadily increased with years. The slope of the sizespectra usually has a negative correlation with fi shing effort, and it has often been used as an indicator to reflect the status of exploited species in other studies(Greenstreet and Hall, 1996; Rice and Gislason, 1996;Gislason and Rice, 1998; Bianchi et al., 2000). Xu and Jin (2005) pointed out that the population dynamics of Japanese anchovy covered the variations in size-spectra of the fi sh community. To remove this inf l uence, the correlation coeffcient of fi shing effort and the slope of the size-spectra was recalculated excluding Japanese anchovy. Still, the results showed no significant differences (Spearman’s rho=0.47,P>0.05,n=10). This inconsistency between the slope of size-spectra and fi shing effort has been found in comparable research (Benoı̂t and Rochet, 2004; Shin and Cury, 2004). To use the slope of size-spectra versus another index as an indicator of fi shing effort merits further investigation.
Although the negative correlation between MIW and years was not significant for some species, theG-test showed that the MIW of the whole fi sh assemblage declined significantly (P<0.05) over time, from 2003 to 2015. This is consistent with the fi ndings of Xu and Jin (2005) for the period 1985-2002. Hence, the trend for decreasing MIW over time has not improved.Furthermore, a significant reduction (P<0.05) in CPUE was observed for 2003-2015 in this study, yet not for 1985-2002 by Xu and Jin (2005). This phenomenon warrants special attention because it occurred not only for some of the traditional economically important species, like largehead hairtail and small yellow croaker, but also for some small-sized and low-value species, like skinnycheek lanternfish and glowbelly. Indicator-based analyses to assess long-term changes in the status of coastal fishes in the Baltic Sea showed a generally ‘deteriorating’status during the past three decades, yet have pointed towards an ‘improving’ status in recent years, which may be due to an improved quality of coastal habitat and/or better controls of fi shing intensity (Bergström et al., 2016). Thus, coordinated local measures may provide a new way to improve the overall status of coastal fishes.
5 CONCLUSION
This work used trawl data to examine fi sh diversity and community structure from 2003 to 2015 in the central and southern Yellow Sea. Fish assemblage indices fluctuated without clear trends from 2003 to 2015, although strong positive and significant correlations were observed among the five indices.Japanese anchovy, anglerfish, Tanaka’s snailfish,silver pomfret, and small yellow croaker were the top-fi ve dominant species during the survey years.There was a high level of similarity (63.71%) among all years. No consistent trends were found in intercept and slope (P>0.05) in size-spectra analysis. There were also no significant differences (P>0.05) between the slope of size-spectra and fi shing effort. Lastly,both the CPUE and MIW analyses of the whole fi sh assemblage showed a significantly decreasing tendency over time (P<0.05).
6 ACKNOWLEDGEMENT
We would like to acknowledge the crew of R/VBeidoufor their efforts in fi eld sampling. We are also grateful to two anonymous reviewers and our colleagues who provided useful comments on the manuscript.
猜你喜欢
杂志排行
Journal of Oceanology and Limnology的其它文章
- Response of the North Pacific Oscillation to global warming in the models of the Intergovernmental Panel on Climate Change Fourth Assessment Report*
- Effect of mesoscale wind stress-SST coupling on the Kuroshio extension jet*
- Surface diurnal warming in the East China Sea derived from satellite remote sensing*
- Cross-shelf transport induced by coastal trapped waves along the coast of East China Sea*
- Observations of near-inertial waves induced by parametric subharmonic instability*
- Seasonal variation and modal content ofinternal tides in the northern South China Sea*