外周血circRNA的差异表达及其与抑郁症严重程度的关系
2018-07-09孔令明牛威姚高峰朱晓丽何明骏张理义
孔令明,牛威,姚高峰,朱晓丽,何明骏,张理义
抑郁症的核心症状是情绪低落、兴趣减退等,重症抑郁障碍者自杀占所有自杀的33%~50%,其共病心血管疾病、糖尿病的机会明显高于正常人,严重威胁着人类的心身健康[1-3]。世界卫生组织预测,到2020年,抑郁症将占据世界疾病负担的第2位[4-5]。探讨抑郁症发病机制及其生物标志物,一直是精神病遗传学领域的研究热点。目前,关于抑郁症的发病机制仍有诸多不明确的方面。有研究者认为,抑郁症可能是由突触可塑性受损致使大脑不能恰当地适应环境所致;还有证据表明,抑郁症与学习和记忆功能受损相关,提示抑郁症存在大脑功能和神经元突触可塑性改变的证据[6-8]。但目前仍然无单一的理论可完全解释抑郁症的发病过程。抑郁症的分子遗传学研究认为,社会环境、不良成长经历可导致某些特定的基因发生表观遗传改变,并引起神经元的可塑性和功能异常。
已有研究证实,circRNA对核酸酶不敏感,因而在精神障碍的发病机制中可起到重要的调节作用[9-10]。circRNA可能参与调控抑郁症发病过程,并与抑郁症的症状表现、严重程度有一定关系[11]。本研究探讨抑郁症患者外周血circRNA的差异表达及其与抑郁症严重程度的关系。
1 资料与方法
1.1 研究对象 2015年4月至2016年10月在解放军102医院门诊及住院的抑郁症患者100例,纳入抑郁症组。入组患者均满足美国精神疾病诊断与统计手册第5版(DSM-Ⅴ)中的MDD诊断标准[12]。诊断由两名精神科医师根SCID-I/P的诊断标准对患者分别进行测评。这两名精神科医师间的评分一致性是0.87,使用HAMD量表中的24条目评估抑郁症患者的严重程度。入组标准:①入组前3个月内未使用抗抑郁药物;②汉族,年龄18~60岁;③无严重器质性疾病史(例如心脏病、糖尿病、帕金森综合征),入组前6个月未发生严重的生活创伤事件;④无其他精神疾病(精神分裂症、躁狂症或人格障碍)。选取同时期健康人100名作为对照。本研究获得中国人民解放军102医院医学伦理委员会批准,所有受试者或受试家属(监护人)均签署知情同意书。
1.2 评估方法 由精神科医师根据DSM-IV-TR对患者进行诊断,如果出现不一致情况,则请一位精神科主任医师共同讨论后确定诊断。在开展研究前,首先对参加本课题的精神专科医师和研究生进行培训。采用汉密尔顿抑郁量表(Hamilton depression scale,HAMD)对患者的抑郁症状进行评估。入组时记录一般情况(姓名、性别、年龄、民族、文化程度、职业、收入水平、婚姻状况、药物成瘾史及精神疾病家族史等)。同时记录对照组一般情况。
1.3 评估工具 HAMD是目前最经典的抑郁他评量表,已广泛应用。该量表主要用于评定个体的抑郁症状严重程度,共有24个条目,7个因子,包括焦虑/躯体化(anxiety/somatization,AS)、体重(weight,W)、认知障碍(cognitive disorder,CD)、日夜变化(diurnal variation,DV)、迟滞(retardation,R)、睡眠障碍(sleep disturbance,SD)和绝望感(hopelessness,H),能够较好地反映患者病情[13]。
1.4 实验过程 实验采用国际标准化流程[14],包括RNA提取、circRNA芯片筛查、RT-PCR验证等。
1.5 统计学处理 采用SPSS 17.0软件工具包进行数据分析。以Spearman相关分析探究差异表达的circRNA与患者HAMD总分及各因子分的相关性。进一步使用Wilcoxon秩和检验比较症状高分组与低分组circRNA表达水平的差异。以logistic回归分析探究circRNAs对抑郁严重程度的贡献,并构建ROC曲线分析相应circRNA对症状严重程度的预测价值。所有统计均为双侧显著性检验,P<0.05为差异有统计学意义。
2 结 果
2.1 抑郁症患者circRNA表达水平与抑郁症症状的相关分析 circRNA_100018的ΔCt值与认知障碍呈明显负相关(P<0.05)。circRNA_104600的ΔCt值与认知障碍呈明显负相关(P<0.05),与睡眠障碍呈明显正相关(P<0.05)。circRNA_103636的ΔCt值与焦虑/躯体化、认知障碍呈明显负相关(P<0.05);circRNA_100679的ΔCt值与体重、绝望感呈明显正相关(P<0.05);circRNA_103964的ΔCt值与焦虑/躯体化呈明显负相关(P<0.05,表1)。
2.2 抑郁症组抑郁症状与circRNA表达水平的回归分析 以HAMD的各因子分为因变量,以circRNA的ΔCt值为自变量行多元回归分析。结果表明,circRNA_100679对抑郁症组体重、绝望感有显著预测作用(P<0.01或P<0.05),对体重、绝望感变异的解释率分别为13.9%、11.1%;circRNA_104121、circRNA_102802和circRNA_104953对迟滞有明显预测作用(P<0.05),可解释迟滞因子变异的16.9%。circRNA_104600、circRNA_103636对睡眠障碍有明显预测作用(P<0.05),可解释睡眠障碍变异的15.4%(表2)。

表1 抑郁症患者circRNA表达水平与抑郁症症状的相关分析(r)Tab.1 Correlation analysis of circRNA expression level and depression symptom in MDD patients (r)

表2 抑郁症组抑郁症状与circRNA表达水平的回归分析Tab.2 Regressive analysis of circRNA expression level and depression symptoms of MDD patients
2.3 抑郁症状不同严重程度circRNA表达水平的比较 以HAMD总分为标准,采用四分位法将抑郁症组分为高分组(前25%)和低分组(后25%)。采用独立样本的Wilcoxon秩和检验比较抑郁高分及低分组circRNA表达水平的差异,发现两组患者hsa_circRNA_002143、circRNA_102802、circRNA_103636, circRNA_100679和circRNA_104953表达水平不同,差异有统计学意义(P<0.05,表3)。2.4 circRNA表达水平对抑郁症患者严重程度的预测价值 以circRNA_002143、circRNA_102802、c i r c R N A_1 0 3 6 3 6、c i r c R N A_1 0 0 6 7 9和circRNA_104953检验变量,以抑郁症组HAMD总分的高和低分组作为状态变量,建构ROC曲线(图1、表4)。结果显示,抑郁症circRNA_100679表达水平对其症状严重程度有中等预测效力(AUC=0.708,P<0.05,95%CI:0.523~0.894)。当血浆中circRNA_100679=17.80685CT值时,约登指数达最大值0.283,灵敏度0.750,特异度0.533。
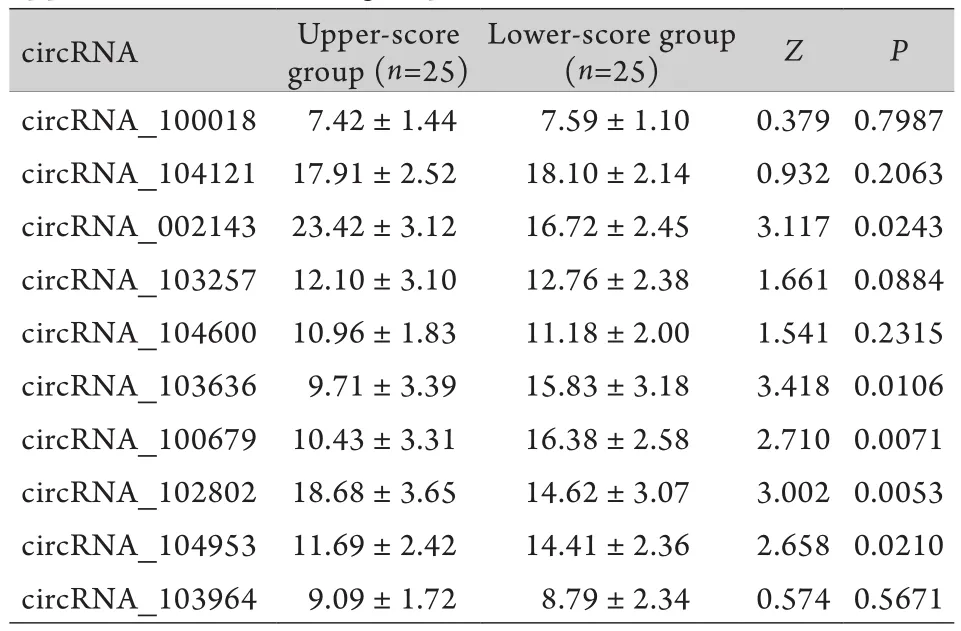
表3 抑郁症状高低分两组circRNA表达水平的比较Tab.3 Comparison of circRNA expression level between upper- and lower-score group of HAMD

图1 circRNA_100679对抑郁症严重程度预测的ROC曲线Fig.1 ROC curve of prediction of circRNA_100679 on severity of depression

表4 抑郁症组HAMD高低分组差异表达circRNA的ROC曲线分析结果Tab.4 Analysis of ROC curve differential expressed of circRNAs
3 讨 论
抑郁症是包含多个症状表现的症状群,且因家庭环境、成长经历、父母教养方式、人格特征等方面呈现个体间变异性。临床实践发现,抑郁症发病率呈现逐年增加的趋势[15-16]。人类对于抑郁症的认知、早期识别、诊断、治疗、康复、复发预防等方面都不能适应现实的需求,尤其是对抑郁症的病理机制和过程仍缺乏充分的了解。
抑郁症症状表现存在较大个体差异[17-18]。本研究对抑郁症状与circRNA表达水平的相关及回归分析发现,circRNA_100018的ΔCt值与认知障碍呈明显负相关;circRNA_104600的ΔCt值与认知障碍呈明显负相关;circRNA_103636的ΔCt值与焦虑/躯体化、认知障碍呈明显负相关;circRNA_103964的ΔCt值与焦虑/躯体化呈明显负相关。circRNA_104121、circRNA_102802和circRNA_104953进入抑郁组迟滞的回归方程,对迟滞因子变异的解释率0.169;circRNA_104600、circRNA_103636进入抑郁组睡眠障碍的回归方程,对睡眠障碍变异的解释率为0.154。说明这些circRNA表达水平越高,抑郁症患者可出现越严重的认知障碍、焦虑/躯体化、迟滞等症状和表现。circRNA_100679的ΔCt值与体重、绝望感呈显著正相关,circRNA_104600的ΔCt值与睡眠障碍也呈正相关,circRNA_100679进入抑郁症组体重、绝望感的回归方程,对体重、绝望感变异的解释率分别为0.139、0.111,提示circRNA表达水平越高,其体重、绝望感、睡眠问题越少。本研究结果发现,circRNA与抑郁症的关系较为复杂,取决于患者的症状表现,而且与circRNA对病理过程的间接调控有关,以往研究发现,circRNA利用其miRNA应答元件结合miRNA,以阻断miRNA对其靶标表达的抑制作用,从而调控其他相关RNA的表达水平[19],进而对疾病的病理过程发挥作用。
抑郁症状高低分组circRNA表达水平的比较发现,抑郁症高分组hsa_circRNA_002143、c i rc R N A_1 0 2 8 0 2表达水平下降;抑郁症高分组circRNA_103636、circRNA_100679和circRNA_104953表达水平升高;进一步以这些circRNAs表达水平为检验变量建构ROC曲线,结果表明,circRNA_100679(AUC=0.708,当血浆中circRNA_100679=17.80685CT值时,约登指数最大值为0.283,灵敏度0.750,特异度0.533)对抑郁症严重程度有较好的预测价值,提示抑郁症病情与circRNA表达水平有密切关系。
circRNA可能在某些疾病病理方面起一定作用,circRNA可通过竞争性结合与疾病关联的miRNA参与疾病发病过程的调节[19-21],例如,环状RNA ciRS-7在人脑组织中丰富表达,与脑特异性microRNA miR-7相互作用;ciRS-7含有多个串联的miR-7结合位点,因此,可以作为内源性的miRNA海绵,抑制miR-7活性;Fan等[10]的研究发现,miRNA-26b、miRNA-1972、miRNA-4485、miRNA-4498和miRNA-4743等可能参与抑郁症的病理过程,这些miRNA的靶基因的合集为734个,涉及的生物学过程包括皮层发育、轴突导向和延伸、突触传递以及学习和记忆过程等,涉及的信号通路包括轴突导向、谷氨酸能突触、Wnt信号通路、ErbB信号通路、mTOR信号通路、VEGF信号通路等,有望成为抑郁症诊断的生物标记物[9,22-26]。circRNA可能通过调控mircRNA的表达水平来影响抑郁症的病理生理过程。
综上所述,本研究发现circRNA_100679对抑郁症严重程度有较好的预测价值;hsa_circRNA_002143、circRNA_100679、circRNA_103636表达水平与患者的抑郁症状有密切关系。
[1] Tolentino JC, Schmidt SL. Association between depression severity and cardiac autonomic modulation[J]. J Psychos Res,2016, 85(6): 9-11.
[2] Patel SS, Mehta V, Changotra H, et al. Depression mediates impaired glucose tolerance and cognitive dysfunction: A neuromodulatory role of rosiglitazone[J]. Horm Behav, 2016,78(2): 200-210.
[3] Hom MA, Lim IC, Stanley IH, et al. Insomnia brings soldiers into mental health treatment, predicts treatment engagement,and outperforms other suicide-related symptoms as a predictor of major depressive episodes[J]. J Psychiatr Res, 2016, 79(8):108-115.
[4] Otte C, Wingenfeld K, Kuehl Linn K, et al. Cognitive function in older adults with major depression: Effects of mineralocorticoid receptor stimulation[J]. J Psychiatr Res, 2015, 69(9): 120-125.
[5] Bradley KAL, Colcombe S, Henderson SE, et al. Neural correlates of self-perceptions in adolescents with major depressive disorder[J]. Dev Cogn Neurosci, 2016, 19(7): 87-97.
[6] Holt RJ, Graham JM, Whitaker KJ, et al. Functional MRI of emotional memory in adolescent depression[J]. Dev Cogn Neurosci, 2016, 19(6): 31-41.
[7] Tao WW, Dong Y, Su Q, et al. Liquiritigenin reverses depressionlike behavior in unpredictable chronic mild stress-induced mice by regulating PI3K/Akt/mTOR mediated BDNF/TrkB pathway[J]. Behav Brain Res, 2016, 308(7): 177-186.
[8] Reimets R, Raud S, Loomets M, et al. Variability in the effect of antidepressants upon Wfs1-deficient mice is dependent on the drugs' mechanism of actions[J]. Behav Brain Res, 2016, 308(7):53-63.
[9] Xia T, Jiang XM, Chen XM, et al. Circular RNA: A new member of competing endogenous RNAs[J]. Chin J Cell Biol, 2013,35(11): 1695–1700. [夏天, 蒋孝明, 陈晓敏, 等. 环状RNA:竞争性内源RNA新成员[J]. 中国细胞生物学学报, 2013,35(11): 1695-1700.]
[10] Fan HM, Sun XY, Guo W, et al. Differential expression of microRNA in peripheral blood mononuclear cells as specific biomarker for major depressive disorder patients[J]. J Psychiatr Res, 2014, 59(11): 45-52.
[11] Kong LM, Niu W, Zhu XL, et al. Correlation of circRNAs'differential expression to negative-positive symptoms of patients with schizophrenia[J]. Med J Chin PLA, 2017, 42(10): 897-901.[孔令明, 牛威, 朱晓丽, 等. 精神分裂症circRNA的差异表达及其与症状的关系[J]. 解放军医学杂志, 2017, 42(10): 897-901.]
[12] Zhang QL, Lu J, Sun XY, et al. A preliminary analysis of association between plasma microRNA expression alteration and symptomatology improvement in major depressive disorder(MDD) patients before and after antidepressant treatment[J].Eur J Psychiat, 2014, 28(4): 252-264.
[13] American Psychiatric Association. Diagnostic and statistical manual of mental disorders (5th ed.)[M]. Washington DC:American Psychiatric Association, 2013. 160-168.
[14] Hamilton M. Development of a rating scale for primary depressive illness[J]. Br J Soc Clin Psychol, 1967, 6(4): 278-296.
[15] Cui X, Niu W, Kong L, et al. hsa_circRNA_103636: potential novel diagnostic and therapeutic biomarker in Major depressive disorder[J]. Biomark Med, 2016, 10(9): 943-952.
[16] Wang XL, Wang DL. Hypothalamic abnormality and major depressive disorder[J]. Adv Psychol Sci, 2015, 23(10): 1763-1774. [王晓乐, 王东林. 下丘脑异常与抑郁症.心理科学进展, 2015, 23(10): 1763-1774.]
[17] Robinson RL, Grabner M, Palli SR, et al. Covariates of depression and high utilizers of healthcare: Impact on resource use and costs[J]. J Psychos Res, 2016, 85(6): 35-43.
[18] Sheehan DV, Nakagome K, Asami Y, et al. Restoring function in major depressive disorder: A system review[J]. J Affect Dis,2017, 215(6): 299-313.
[19] Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency[J]. Nature, 2013,495(7441): 333-338.
[20] Wilusz JE, Sharp PA. Molecular biology. A circuitous route to noncoding RNA[J]. Sci, 2013, 340(6131): 440-441.
[21] Jeck WR, Sorrentino JA, Wang K, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats[J]. RNA,2013, 19(2): 141-157.
[22] Vandeleur CL, Fassassi S, Castelao E, et al. Prevalence and correlates of DSM-5 major depressive disorder and related disorders in the community[J]. Psychiatr Rese, 2017, 250(1):50-58.
[23] Memczak S, Jens M, Elefsinioti A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency[J]. Nature, 2013,495(7441): 333-338.
[24] Hansen TB, Jensen TI, Clausen BH, et al. Natural RNA circles function as efficient microRNA sponges[J]. Nature, 2013,495(7441): 384-388.
[25] Bingol B, Sheng M. Deconstruction for reconstruction: the role of proteolysis in neural plasticity and disease[J]. Neuron, 2011,69(1): 22-32.
[26] Fan HM, Wu WB, Niu W, et al. Bioformatics analysis of microRNA differentially expressed in major depressive disorder[J]. Chin J Behav Brain Sci, 2015, 24(5): 418-422. [范惠民, 吴文波, 牛威, 等. 重症抑郁障碍外周血microRNA的生物信息学分析[J]. 中华行为医学与脑科学杂志, 2015, 24(5):418-422.]
