小麦冬前次生根对生育中后期地上部发育及产量的影响
2018-07-06杨习文周苏玫徐凤丹梅晶晶申冠宇李秋杰贺德先
杨 蕊,杨习文,周苏玫,周 燕,徐凤丹,梅晶晶,申冠宇,李秋杰,贺德先
(河南农业大学 农学院,河南粮食作物协同创新中心,小麦玉米作物学国家重点实验室,河南 郑州 450002)
发达的根系有助于小麦(TriticumaestivumL.)植株健壮生长和产量提高[1],其中,次生根是小麦根系的主要组成部分,主要分布在养分丰富的耕作层,对于水分和养分吸收能力强[2-3]。另外,次生根发生时期长、数目多、根间补偿作用明显,对巩固分蘖,促进穗部发育,提高籽粒灌浆和结实率具有决定性作用[4]。次生根生长有2个旺盛时期,一是冬前分蘖期,二是春季分蘖期[5]。冬前分蘖期发根数占全生育期次生根发根总数的35%,其数目的多少一方面是衡量壮弱苗的标准,另一方面对植株的生长发育也有重要影响,因此,研究冬前单株次生根对小麦生育中后期植株生长发育的影响显得尤为重要。采取相应的耕作与栽培技术措施,以充分挖掘次生根在籽粒增产中的作用,是小麦高产栽培的一个重要途径。小麦不同品种在相同栽培条件,或同一品种在不同栽培条件下,其单株次生根数均有可能出现明显的差异;相同品种在相同栽培条件下,不同植株单株次生根也可能存在一定的差异。次生根有利于巩固和发展小麦根系,促进地上部健壮生长,这对实现高产稳产具有重要作用[6-9]。薛丽华等[10]研究调亏灌溉下次生根对小麦生长的影响,发现拔节期减少次生根数会显著降低小麦的粒质量。越冬前次生根数与成穗数关系密切。而方燕等[11]发现在越冬期适当减少根量(多为次生根)显著增加了小麦的产量。李鸿斐等[12]发现次生根数与籽粒品质有关。刘殿英等[5]研究指出,小麦单株次生根数与分蘖数、穗数、光合强度和地上干质量呈正相关。次生根数对产量以及品质的影响不容忽视,因此,研究次生根数对植株生育中后期及籽粒产量的影响,具有一定的理论和实践意义。不同品种以及不同栽培条件对次生根的影响较大,前人研究也较多,但试验过程中操作难度较大,导致精确度低,操作不当可能会对植株生长发育造成较大影响,且结论也存在差异。而研究冬前次生根对植株的生长发育和产量形成的影响,量化次生根的作用未见报道。鉴于此,本试验采用大田育苗、越冬前根据次生根数分组移栽的方式,在无损伤或尽可能减少根系损伤的条件下,明确越冬前次生根数量对小麦植株地上部发育及籽粒产量的影响,以期为有针对性地调控根系发育,优化根系构型提供参考,从而达到提高产量的目的。
1 材料和方法
1.1 供试材料与试验设计
试验于2015年10月-2016年6月在河南农业大学科教园区(郑州市惠济区毛庄镇)进行,供试土壤有机质、全氮、碱解氮、有效磷和有效钾含量分别为17.8 g/kg,0.89 g/kg,57.9 mg/kg,24.4 mg/kg,154.8 mg/kg;2016年10月-2017年6月在河南省农业高新科技园(郑州市中牟县官渡镇)进行,供试土壤有机质、全氮、碱解氮、有效磷和有效钾含量分别是11.3 g/kg,0.75 g/kg,49.8 mg/kg,19.8 mg/kg,134.5 mg/kg。
施肥情况:育苗区纯氮、P2O5和K2O总施用量分别为240,135,120 kg/hm2,全部施入;盆栽纯氮、P2O5和K2O总施用量分别为720,135,120 kg/hm2,其中全部磷肥和钾肥及50%氮肥基施,另外50%氮肥于拔节期随灌水追施,其他管理同一般高产田。
供试小麦品种为矮抗58,分别于2015年10月15日和2016年10月19日播种,于2015年12月15日和2016年12月17日越冬前根据单株次生根数分组移栽入盆。盆栽所用花盆高30 cm,直径25 cm,每盆装大田耕层土壤10 kg,并均匀混入肥料。具体次生根分组情况:选取单株总分蘖数为4(包括主茎)的幼苗,将植株根系完整挖出,根据单株次生根数分为:处理A(次生根数较少,次生根1~2条)、处理B(次生根数中等,次生根3~4条)和处理C(次生根数较多,次生根5条及以上),共计3个处理,每盆定植10株,每个处理重复24盆。
1.2 测定项目及方法
小麦生长发育期间,2年分别于返青期(RS表示:2016年2月10日、2017年2月10日),拔节期(JS表示:2016年3月19日、2017年3月17日),抽穗期(HS表示:2016年4月14日、2017年4月12日),灌浆期(GS表示:2016年5月15日、2017年5月12日),蜡熟期(LS表示:2016年5月25日、2017年5月25日)取样。在返青期、拔节期、抽穗期、灌浆期、蜡熟期,测定单株蘖数、地上部干物质量、株高、单株根系生理势;在返青期、拔节期、抽穗期、灌浆期,测定单株叶面积、叶片叶绿素含量(SPAD值)。测定方法如下:
单株蘖数:每个处理选取有代表性的植株10株,确定存活的单株蘖数。
单株叶面积:采用比叶重法测定[13]。
株高:蜡熟期前测定分蘖节到最上展开叶叶尖的高度,蜡熟期测定分蘖节到最高穗穗尖的高度。
叶片叶绿素含量(SPAD值):叶绿素测定仪测得的叶片SPAD值为叶绿素含量的相对值,原理是利用叶片对不同波长光吸收的差异来估算叶绿素含量[14-15]。采用SPAD-502 Plus叶绿素测定仪测定,测定部位为最上展开叶的中部,每个处理重复5次。
地上部干物质量:每个处理选取5株有代表性的植株,去除根系后在105 ℃条件下杀青15 min,然后在80 ℃条件下烘至恒质量(24 h),用1‰电子天平称取每株的质量。
单株根鲜质量:每个处理选取3株有代表性的植株,去除地上部后,将根系清洗干净后在滤纸上吸去多余的水分,用1‰电子天平称质量(g)。
根系活力测定:采用改良TTC法,参考《植物生理学实验指导》[16],每个处理重复3次。
单株根系生理势:单株根鲜质量与根系活性的乘积,单位μg/h。
成熟期取6个重复计产,同时调查产量构成因素。
1.3 数据分析方法
使用Microsoft Excel 2010和SPSS 19.0进行数据处理和图表制作,并通过Duncan法进行方差分析。
2 结果与分析
2.1 小麦冬前次生根数对单株蘖数的影响
2年试验结果表现出一致趋势(图1)。小麦生育中后期不同处理单株蘖数均呈先升后降趋势,抽穗期达到最大;各个生育期单株蘖数随冬前单株次生根数增加呈递增趋势。2015-2016年,在返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C的单株蘖数比处理A分别增加了26.7%,28.6%,50.0%,6.3%,7.7%,返青期、拔节期、抽穗期差异达显著水平;在返青期、拔节期和抽穗期,处理C比处理B增加了18.8%,12.5%,20.0%,差异均达显著水平,而处理A和处理B的单株蘖数差异不显著。2016-2017年,在返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C比处理A分别增加了26.3%,13.0%,85.7%,13.0%,30.0%,其中,返青期和抽穗期差异达显著水平,处理B比处理A分别增加了5.3%,8.7%,57.1%,8.7%,20.0%,抽穗期差异达显著水平。由此可见,增加冬前单株次生根数可以显著提高生育中后期植株的单株蘖数。

A.次生根数较少;B.次生根数中等;C.次生根数较多。图柱上的不同字母表示同一时期不同处理间差异显著(P<0.05)。图2-3同。A.Less nodal roots per plant;B.Medium nodal roots per plant;C.More nodal roots per plant. Different letters above the columns indicated significant difference among the different treatments in the same period at 0.05 level. The same as Fig.2-3.
2.2 小麦冬前次生根数对地上部干物质量的影响
随着生育期推进,2年不同处理地上部干物质量呈现逐渐增加的趋势;随冬前单株次生根数增加不同处理间地上部干物质量均随之增加,表现为处理A <处理B <处理C(图2)。随生育进程推进,次生根数对地上部干物质量的影响在不同处理间的差异逐渐减小。2015-2016年,在返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C比处理A地上部干物质量分别提高了25.5%,39.4%,37.6%,14.5%,11.7%,处理C比处理B分别提高了10.5%,23.5%,30.7%,10.2%,3.8%,其中,返青期和拔节期3个处理间差异达显著水平,抽穗期处理C与处理A、B差异显著,但处理A和B差异不显著;2016-2017年,在返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C的地上部干物质量比处理A分别增加了23.5%,28.3%,30.5%,27.4%,21.0%,处理C比处理B分别增加了6.2%,28.0%,2.3%,26.6%,19.1%,其中,返青期处理A与处理B、C差异达显著水平,但处理B和C差异不显著。表明适当增加冬前单株次生根数可提高生育中期植株地上部干物质量,对生育后期影响相对较小。

图2 小麦冬前次生根数对植株地上部干物质量的影响Fig.2 Effect of prior-to-winter nodal roots to dry matter on the ground
2.3 小麦冬前次生根数对株高和单株叶面积的影响
随着小麦生育进程的推进,综合2年结果,不同处理株高均呈先升后降的变化趋势;在返青期、抽穗期、蜡熟期,不同处理间2年差异未达显著水平(表1)。从2年平均值来看,在拔节期、抽穗期、蜡熟期,处理C的株高比处理A分别增加13.9%,0.5%,4.0%,处理C比处理B分别增加15.3%,1.2%,3.1%,其中,仅在拔节期处理C与处理A、B差异达显著水平,而在返青期和灌浆中期,处理C的株高低于处理A、B。由此表明,越冬前单株次生根数对植株生育中后期株高有一定的影响,但整体上影响相对较小。

表1 小麦冬前次生根数对株高的影响Tab.1 Effect of prior-to-winter nodal roots to the height of plant cm
从表2可以看出,随生育进程推进,小麦单株叶面积呈缓慢上升的趋势,在灌浆中期达到最大值,由于蜡熟期小麦叶片干枯破损严重,未进行叶面积测定;在返青期、拔节期、抽穗期、灌浆中期,随冬前单株次生根数增加小麦单株叶面积有增加的趋势。从2年平均值来看,处理C比处理A在返青期、拔节期、抽穗期、灌浆中期分别增加了43.7%,14.0%,8.3%,9.4%,差异均达显著水平;处理C比处理B在返青期、拔节期、抽穗期、灌浆中期分别增加了15.3%,-3.1%,16.4%,29.2%,其中,抽穗期和灌浆中期差异达显著水平。这说明越冬前单株次生根数增加可提高生育中后期单株叶面积。
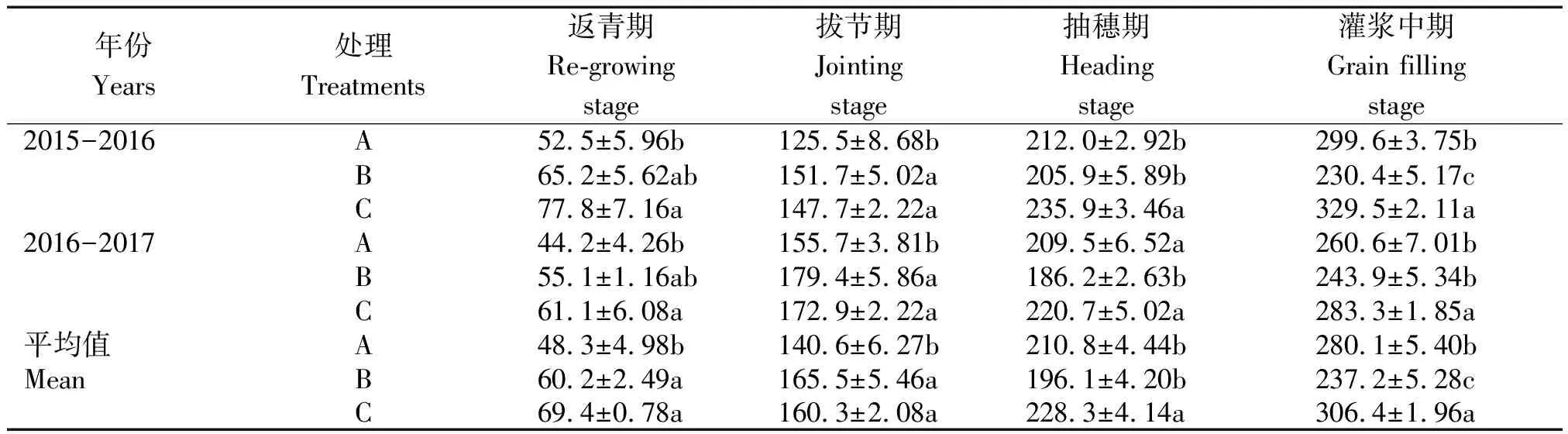
表2 小麦冬前次生根数对单株叶面积的影响Tab.2 Effect of prior-to-winter nodal roots to leaf area per plant cm2
2.4 小麦冬前次生根数对叶片叶绿素含量(SPAD值)的影响
在本试验条件下,由于蜡熟期小麦叶片干枯破损严重,未进行叶片叶绿素含量的测定,返青期-灌浆期不同处理叶片叶绿素含量(SPAD值)的变化为35.1~59.7,随着生育期推进,同一处理的叶片SPAD值基本呈先升后降再升的变化趋势,不同处理对生育中后期旗叶SPAD值的影响较小(表3),但存在随着次生根数增加,叶片SPAD增加的趋势。从2年的平均值来看,在返青期、拔节期、抽穗期、灌浆期,处理C比处理A旗叶SPAD值分别增加了2.6%,2.3%,5.4%,-0.7%,处理B比处理A分别增加了7.2%,-1.1%,8.2%,-4.8%,处理C比处理B分别增加了-4.3%,3.4%,-2.6%,4.3%,其中,返青期和抽穗期处理A与处理B、C差异达显著水平,但处理B和C差异不显著。由此说明,冬前单株次生根数对旗叶叶绿素含量的影响虽然较小,但存在随次生根数增加,旗叶叶绿素含量增加的趋势。

表3 小麦冬前次生根数对叶片SPAD值的影响Tab.3 Effect of prior-to-winter nodal roots to the SPAD of leaves
2.5 小麦冬前次生根数对单株根系生理势的影响
不同处理单株根系生理势整体变化趋势表现为先升后降,2年均于拔节期达到最大;不同处理根系生理势随冬前次生根数的增加而增加(图3)。在2015-2016年返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C的单株根系生理势比处理A分别增加了106.7%,91.2%,83.5%,58.6%,244.5%,其中,返青期、拔节期、灌浆中期和蜡熟期差异达显著水平,处理C比处理B分别增加了79.8%,62.0%,48.6%,40.6%,40.6%,在返青期、拔节期和蜡熟期差异达显著;2016-2017年的返青期、拔节期、抽穗期、灌浆中期、蜡熟期,处理C单株根系生理势比处理A分别增加了515.0%,237.1%,25.5%,79.9%,40.9%,其中,各个测定时期差异均达显著水平,处理C比处理B分别增加了42.8%,44.1%,10.2%,68.4%,5.2%,仅在拔节期和灌浆中期差异达显著水平。因此,冬前单株次生根数较多有利于单株根系生理势的提高,使根系维持较高的活性。

图3 小麦冬前次生根数对单株根系生理势的影响Fig.3 Effect of prior-to-winter nodal roots to physiological potential of per plant roots
2.6 小麦冬前次生根对籽粒产量及其构成因素的影响
从表4可以看出,单株产量2年均表现出处理C>处理B>处理A,处理C的产量比处理A、B,分别增加了52.4%,33.3%,差异达显著水平。从2年平均值来看,随冬前次生根数增加,不同处理间单株穗数同样表现出:处理 C>处理B>处理A,处理C的单株穗数比处理A、B分别增加了25.0%,25.0%,差异达显著水平;结实小穗数也随冬前次生根数增加而呈递增趋势,处理C比处理A、B增加了13.4%,4.4%;从2年平均值来看,不孕小穗数表现出处理B>处理C>处理A,处理C和A差异未达显著水平,处理A、C比处理B的不孕小穗数降低了37.5%,30.0%。表4还表明,随冬前次生根数增加穗粒数有所增加,处理C比处理A、B分别增加26.2%,12.0%,处理B比处理A增加12.6%,3个处理差异均达显著水平。由此表明,小麦冬前单株次生根数增加,有利于增加单株穗数、结实小穗数和穗粒数,降低不孕小穗数,进而提高产量。

表4 小麦冬前次生根数对籽粒产量及其构成因素的影响Tab.4 Effect of prior-to-winter nodal roots to grain yield and its components
3 结论与讨论
冬前次生根影响生育中后期植株地上部的生长发育、根系功能及籽粒产量。小麦冬前分蘖期是促根增蘖、培育壮苗、保证植株安全越冬的关键时期[17]。次生根发育的好坏是麦苗壮弱的重要标志,但因耕作制度和土壤墒情不同,最适冬前次生根数不尽相同。有研究表明,在冬水前移条件下,小麦冬前次生根数增加,从而延长了旗叶的功能期,使成穗数和千粒质量增加,提高了籽粒产量[18]。另有研究表明,适当减少次生根数,有利于提高小麦生育后期叶片的光合速率和花后旗叶SPAD值[19-20]。本研究表明,增加冬前次生根数可以显著提高小麦生育中后期单株蘖数、地上部干物质量和单株叶面积,但对株高和旗叶SPAD值的影响较小。本试验结果与前人研究结果存在着一些差异,究其原因,可能与小麦品种和种植方式的不同有关,也可能与气候条件和土壤墒情密切相关[21]。小麦生育后期叶片光合产物对籽粒的贡献率可达80%[22],本研究表明,冬前单株次生根数增加促进了地上部植株生长,增加了光合面积,促进生育后期光合产物的形成,这是籽粒产量形成的基础。
前人研究指出,植株的生长发育是地上和地下部分协调发展的结果,地上部可为根系提供充足的光合产物,有利于根系形态结构的建成和生理功能的维持[23]。张素瑜等[24]研究表明,轻旱处理后,小麦单株次生根数增加,由此促进了地上部的生长发育,进而提高了根系活力。本研究中,冬前较多的单株次生根数能显著提高植株的根系生物量,维持较高的根系活性,从而提高了单株根系生理势,延缓了生育后期根系衰老。可见,冬前次生根在扩大根系吸收面积和增强根系生理功能等方面具有极其重要的作用。
在小麦生育中后期,次生根对矿物质和水分吸收起重要作用[25],其生长发育状况对植株地上部生长发育和产量形成有重要影响[26-27]。关于次生根的生长发育及功能,前人已做了多方面的研究,在不同试验条件下,次生根对产量的贡献不同[28-30]。从本研究的产量构成看,增加冬前单株次生根数显著提高了小麦单株穗数、结实小穗数和穗粒数,进而提高了籽粒产量。有研究表明,小麦次生根数与千粒质量存在一定的潜在联系[31-33],本研究条件下次生根对千粒质量无显著影响,但明显影响了单株产量,可能与研究方式不同有关。
综上所述,冬前次生根数增加,提高了生育中后期植株的光合面积,维持了较高的根系吸收能力,进而促进了地上部干物质积累,提高了小麦产量构成因素,从而提高了籽粒产量。因此,在生产实践中,创造有利条件培育壮苗,以保证越冬前有较多的次生根数,是植株健壮生长和提高籽粒产量的基础。但冬前次生根数过多又会造成根系冗余,使大量养分消耗于营养器官的生长。因此,冬前最适宜的单株次生根数还有待于进一步研究探讨。
参考文献:
[1] 张永清. 谷物作物根系生长与调控研究[M]. 北京:中国农业科学技术出版社,2006:246-247.
[2] 马元喜. 小麦的根[M]. 北京:中国农业出版社,1999:23-24.
[3] 宋欣欣,贺德先.小麦生育后期主茎和分蘖次生根对籽粒产量和品质的影响[J]. 麦类作物学报,2011,31(2):281-285.
[4] 董文华. 施氮量对不同种植密度下冬小麦群体结构及产量的影响[D]. 泰安:山东农业大学,2017.
[5] 刘殿英,石立岩,黄炳茹,等. 小麦的根与高效栽培[M]//中国小麦栽培研究新进展. 北京:中国农业出版社,1993:480-490.
[6] 臧贺藏. 河南主栽小麦品种根系性状聚类分析及初生根与次生根发育差异研究[D]. 郑州:河南农业大学,2009:50-51.
[7] 杨荣光,于春霞,张兴强,等. 不同土壤水分条件下冬小麦根系分布规律研究[J]. 耕作与栽培,2014(1):4-5.
[8] 吴永成,周顺利,王志敏. 小麦与抗旱性有关的根系遗传改良研究进展[J]. 麦类作物学报,2004,24(3):101-104.
[9] 马冬云,郭天财,朱云集,等. 不同穗型冬小麦品种根系时空分布特征及其碳氮代谢的研究[J]. 西北植物学报,2006,26(4):778-783.
[10] 薛丽华,段俊杰,王志敏,等. 冬小麦调亏灌溉下次生根和初生根对植株生长的影响[J]. 华北农学报,2010,25(3):87-91.
[11] 方 燕,徐炳成,谷艳杰,等. 密度和修剪对冬小麦根系时空分布和产量的影响[J]. 生态学报,2015,35(6):1820-1828.
[12] 李鸿斐,胡廷积,王晨阳,等. 不同品质类型冬小麦根系基本性状的比较研究[J]. 河南农业大学学报,2001,35(2):107-110.
[13] 王志伟,乔祥梅,程加省,等. 不同小麦品种叶面积、叶绿素相对质量分数、根系性状及产量的研究[J]. 西南大学学报:自然科学版,2016,38(8):10-15.
[14] 张 燕,王百群,何瑞清. 不同施肥下冬小麦生长过程中土壤矿质氮变化及其与冬小麦叶片SPAD值的关系[J]. 水土保持研究,2016,23(6):78-82.
[15] 史力超. 基于叶片SPAD值的滴灌春小麦氮肥分期推荐研究[D]. 石河子:石河子大学,2016.
[16] 刘 萍,李明军,丁义峰. 植物生理学实验[M].北京:科学出版社,2016.
[17] 徐兆飞. 山西小麦[M]. 北京:中国农业出版社,2006:324-341.
[18] 闫翠萍,肖俊红,张 晶,等. 冬水前移对冬小麦生长及水分利用效率的影响[J]. 中国生态农业学报,2018,26(5):677-683.
[19] Berry P M,Spink J H,Gay A P,et al. A comparison of root and stem lodging risks among winter wheat cultivars[J]. The Journal of Agricultural Science,2003,141(12):191-202.
[20] 马守成,徐炳成,李凤民,等. 根系修剪对黄土旱塬冬小麦根系分布,根系效率及产量形成的影响[J]. 生态学报,2008,28(12):6172-6179.
[21] 张 晶,张定一,王 丽,等. 不同有机肥和氮磷组合对旱地小麦的增产机理研究[J]. 植物营养与肥料学报,2017,23(1):238-243.
[22] 刘雪莲,张 洋,张文英,等. 不同基因型冬小麦花后干物质的累积、转运及氮动态变化研究[J]. 青海大学学报:自然科学版,2014,32(5):70-75.
[23] 马富举,李丹丹,蔡 剑,等. 干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响[J]. 应用生态学报,2012,23(3):724-730.
[24] 张素瑜,王和洲,杨明达,等. 水分与玉米秸秆还田对小麦根系生长和水分利用效率的影响[J]. 中国农业科学,2016,49(13):2484-2496.
[25] 左广胜,郑彦周,李雪梅,等. 禾顶囊壳对小麦初生根和次生根的侵染性研究[J]. 中国农学通报,2009,25(19):209-212.
[26] 宋欣欣. 小麦主茎和分蘖根系发育的差异及对籽粒产量和品质的贡献[D]. 郑州:河南农业大学,2011:36-37.
[27] 张 楠. 陕西旱地不同基因型冬小麦生理特性与生产能力的研究[D]. 杨凌:西北农林科技大学,2014.
[28] 张伟杨,钱希旸,李银银,等. 土壤干旱对小麦生理性状和产量的影响[J]. 麦类作物学报,2016,36(4):491-500.
[29] 陈金平,尹志刚,周国勤,等. 不同生育时期浇水对小麦生长发育和产量的影响[J]. 河南农业科学,2016,45(3):39-42.
[30] 王升星,朱玉磊,刘 鹏,等. 小麦次生根数相关分子标记的挖掘[J]. 麦类作物学报,2014,34(12):1627-1632.
[31] 张文渊. 典型生理性状与形态解剖性状对旱地小麦产量形成的差别贡献及进化特征研究[D]. 兰州:兰州大学,2017.
[32] 李海燕,苏 媛,齐永志,等. 多功能土壤添加剂对小麦土传病害的防效及对玉米秸秆的腐解作用[J]. 河南农业科学,2015,44(6):84-89.
[33] 臧贺藏,贺德先.河南省主导小麦品种根系性状的聚类分析[J].河南农业科学,2009,38(5):24-28.
