灌水处理对黄冠梨叶片结构和激素含量及PbDREB基因表达的影响
2018-07-04黄海娜杨金宇李红旭刘小勇叶振风朱立武衡伟
黄海娜,杨金宇,李红旭,刘小勇,叶振风,朱立武,衡伟
(1. 安徽农业大学园艺学院,安徽 合肥 230036; 2. 甘肃省农业科学院果树研究所,甘肃 兰州 730070)
梨(PyrusL.)属于蔷薇科(Rosaceae)、梨亚科(Pomaceae)、落叶乔木[1]。黄冠梨(Pyrusbretschneideri×pyrifolia)是中国培育的优质、丰产早中熟品种,在中国已大量栽培[2],已成为许多省市的支柱产业[3]。中国是一个水资源短缺的国家,尤其是西北大部分地区为干旱、半干旱气候,降雨量少,蒸发量大[4]。因此,研究一种最节水、最有效的灌溉方法对干旱、半干旱地区梨产业的发展具有极其重要作用。覆膜滴灌是地膜覆盖栽培与滴灌技术相结合的高效节水灌溉方式,能有效改善土壤水、热、气、肥条件,从而达到提高产量、改善品质的目的[5]。李昭楠等[6]的研究表明,在甘肃河西走廊干旱荒漠区,膜下滴灌葡萄产量和水分利用效率均高于未覆膜滴灌的产量。李敏敏等[7]对苹果研究发现,膜下滴灌可以提高苹果的产量和水分利用率。
多年来,国内外对果树抗旱机制的研究主要集中在叶片组织结构特征、生理生化以及抗旱基因表达等方面[8]。研究表明,植物的抗旱性与植物叶片的形态解剖结构呈一定的相关性[9-11]。在生理生化与抗旱性关系的研究方面,植物内源激素是对干旱胁迫最为敏感的生理活性物质[12]。随着生物技术的提高,人们对抗旱机制的研究更加深入,脱水反应元件结合蛋白(dehydration responsive element binding, DREB)是一类参与应答植物低温、高盐和干旱等非生物胁迫的主要调控蛋白,特异识别并结合 DRE顺式作用元件,启动下游基因的表达[13]。DREB受低温、高盐和干旱等逆境胁迫的诱导,通过在植物体内的调控表达,从而维持植物细胞的正常生理生化代谢活动,主要表现为细胞膜结构的稳定、细胞渗透压的调节、消除活性氧对细胞的毒害等,提高植物的抗逆性[14]。
本研究以不同的灌水方式浇灌梨园,针对不同灌溉处理对黄冠梨叶片组织结构、激素含量及DREB基因表达量的影响进行研究,以期找出可满足果树高产和提高水分利用率的最佳灌水方式,为该地区梨园节水滴灌提供理论依据和实践指导。
1 材料与方法
1.1 试验材料
试验在兰州市景泰县条山集团梨园内进行,供试品种为生长健壮、均匀一致的‘黄冠’梨,砧木为杜梨,树龄9a。待果实采收后分别从各处理梨树冠外围东、南、西、北面及内膛当年生枝条上随机釆取叶片各30个,置入冰盒带回实验室进行生理指标测定,随机挑取嫩叶进行RNA的提取。
1.2 试验处理
梨树行内覆盖黑色地膜,覆膜宽度2.0 m,植株两边距离树干1.5 m处各设置一条“小孔回流”滴灌带。各处理选择正常植株6株,处理间留2株间隔植株,组合小区,顺序排列,4次重复灌溉水通过水泵抽取,灌水量由水表自动控制流量。覆膜滴灌下渗深度观测,均通过各处理行间预设的100cm壕沟固定观察点进行。实验设6个处理,其中以常规漫灌为对照,具体灌水方式如下:
CK:常规漫灌,分别于花后、果实迅速膨大期和果实膨大期进行灌水,共3次/年,每次灌水量相同,年灌水总量为10200 m3·hm-2。
T1:覆膜滴灌,分别于花前、花后、果实迅速膨大期和果实膨大期(2次)进行灌水,共5次/年,每次下渗60 cm,年灌水总量为4800 m3·hm-2;
T2:覆膜滴灌,分别于花前、花后、幼果期、果实迅速膨大期和果实膨大期(2次)进行灌水,共6次/年,每次下渗60 cm,年灌水总量为5400 m3·hm-2;
T3:覆膜滴灌,分别于花前、花后、幼果期、果实迅速膨大期和果实膨大期(4次)进行灌水,共8次/年,每次下渗40 cm,年灌水总量为5400 m3·hm-2;
T4:覆膜滴灌,分别于花前、花后、幼果期、果实迅速膨大期和果实膨大期(4次)进行灌水,共8次/年,每次下渗60 cm,年灌水总量为6000 m3·hm-2;
T5:覆膜滴灌,分别于花前、花后、幼果期、果实迅速膨大期和果实膨大期(5次)进行灌水,共9次/年,每次下渗60 cm,年灌水总量为6750 m3·hm-2;
1.3 方法
1.3.1 叶片组织结构观察 9月中旬采集自然状态下正常的6种不同灌溉处理梨树叶片,剪取中部成熟叶片主脉中间0.5 cm×0.5 cm的小块,用FAA固定液固定后用石蜡切片法[15]切片观察叶片组织结构。切片厚度8~10 μm,番红-固绿染色,加拿大树胶封片。用显微测微尺测量其叶片总厚度、栅栏组织厚度、海绵组织厚度,每片观察3个视野,重复 6次,每种砧木观察视野总数18个。按以下公式计算叶片组织结构紧密度(CTR)和组织结构疏松度(SR)[11]。
CTR(%)=栅栏组织厚度/叶片厚度×100
SR(%)=海绵组织厚度/叶片厚度×100
1.3.2 激素含量测定 叶片中总赤霉素(GA)、玉米素核苷(ZR)、生长素(IAA)、脱落酸(ABA)等含量测定采用酶联免疫法[16]。
1.3.3 黄冠梨PbDREB基因克隆 利用艾德莱RNA试剂盒(北京艾德莱生物科技有限公司)提取黄冠梨叶片的总RNA,并用1%琼脂糖电泳检测其完整性。用M-MLV反转录酶(Promega公司)合成第一链cDNA,具体反应体系和操作步骤依据说明书进行。
根据其它物种DREB基因的保守序列设计特异性引物,以‘黄冠’梨叶片的cDNA为模板,经PCR扩增目的片段,PCR扩增产物经1%琼脂糖电泳分离后回收,连接到pGEM-T载体中,挑取单菌落进行测序。PCR反应体系为20 μL,反应程序为:94℃预变性3 min,94℃变性30 s,50℃退火30 s,72℃延伸1 min,共30个循环。参照TaKaRa公司的3′-Full RACE Kit试剂盒,根据所获得的目的片段设计3′端引物,经PCR扩增出3′端的片段序列,测序后与目的片段拼接。将所得序列与我们构建的梨EST库比对,找出保守序列设计5′端引物,经PCR扩增出5′端序列,测序后与目的片段拼接。根据拼接序列,设计全长引物,验证拼接结果。试验中所用引物序列如表1所示。

表1 梨PbDREB 基因克隆和表达的相关引物
1.3.4PbDREB基因生物信息学分析 利用相关生物信息学网站对已得基因全长进行生物信息学分析。运用NCBI网站(http://www.ncbi.nlm.nih.gov/)查找和翻译开放阅读框(Open Reading Frame, ORF);运用NCBI数据库中的BLASTX对氨基酸序列进行同源性搜索和比对;运用Protparam(http://www.expasy.ch/tools/protparam.html)对蛋白质理化性质进行在线分析;用TMHMM-2.0(http://www.cbs.dtu.dk/services/TMHMM)对PbDREB氨基酸序列进行分析;运用Net Phos2.0(http://www.cbs.dtu.dk/services/NetPhos/)进行磷酸化位点分析;运用Psort(http://psort.hgc.jp/form.html)进行亚细胞定位分析;氨基酸多重序列比较采用Clustalw2(http://www.ebi.ac.uk/tools/msa/clustalw2/)。
1.3.5PbDREB基因表达分析 分别以黄冠梨叶片的cDNA为模板,以黄冠梨的Actin片段为内参,采用RT-PCR对其进行组织特异性表达分析。根据RACE获得的PbDREB全长序列,使用primer premier 5.0设计荧光定量PCR引物(表1)。使用ABI STEPONE荧光定量PCR仪进行荧光定量PCR分析,采用20 μL反应体系:SYBRTM Premix Ex Taq 10 μL,正、反向引物(10 μmol · L-1)各0.8 μL,ROX Reference Dye 50X 0.4 μL,cDNA模板2.0 μL,ddH2O 6.0 μL。反应程序为:95℃预变性90 s;PCR反应:95℃变性15 s,60℃退火40 s,总共40个循环,每个样品进行4次重复检测,使用2-△△Ct法,求得待测样品相对表达量。
1.4 数据分析
采用 Excel 2003整理实验数据,运用 SPSS 17.0统计软件进行单因素方差分析。
2 结果与分析
2.1 叶片解剖结构特征比较
从叶片的横切解剖结构看(见图1),上下表皮细胞均为单层。5个处理和对照叶片的栅栏组织细胞均为长柱型,排列整齐而紧密,厚度在65.28~82.72 μm之间。海绵组织位于栅栏组织与下表皮之间,形状不规整,排列疏松,多空隙,厚度在101.91~125.41 μm之间,不同处理之间厚度有较大的差异。
由表2可知,与对照相比,5种处理的叶片厚度均显著增大。其中T3的叶片厚度最大,达到257.13 μm;T3、T4和T5的角质层厚度高于对照,T1和T2低于对照;除T2的栅栏组织厚度小于对照外,其它处理均大于对照;除T5的海绵组织厚度与对照差异不显著外,其它处理均显著高于对照;T2的CTR值低于对照,其它处理均高于对照,其中T1最高,T5次之;T1、T2和T4的SR值显著高于对照,T3和T5低于对照,其中T5的SR值最小,但未达到差异显著水平。同时,年灌水量相同(年灌水总量为5400 m3·hm-2)、灌水次数与下渗深度不同的T2与T3比较,T3处理叶片厚度、角质层厚度、栅栏组织厚度、栅栏组织/海绵组织与叶片结构紧密度均明显高于T2,且差异达到显著水平。

图1 不同灌溉处理的黄冠梨叶片横切解剖结构图Fig. 1 Leaves crosscutting anatomical structure pictures of ‘Huangguan’ pear of different irrigation treatments
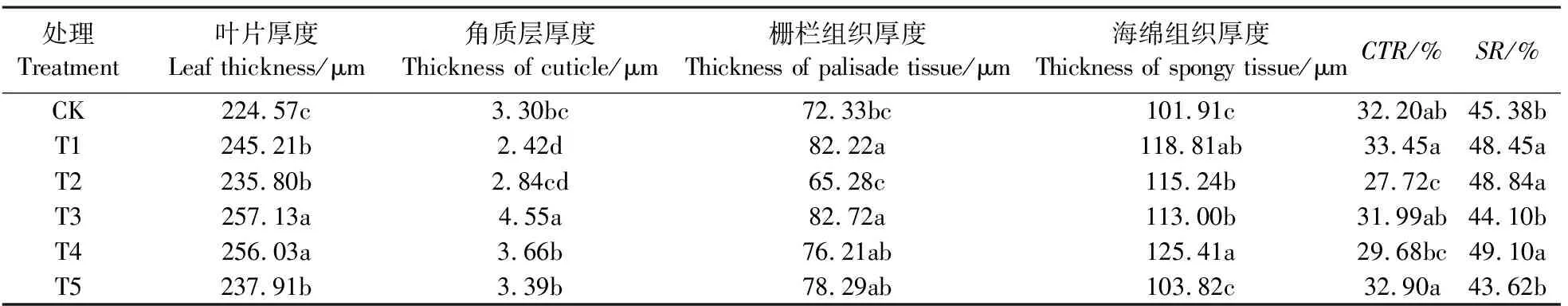
处理Treatment叶片厚度Leaf thickness/μm角质层厚度Thickness of cuticle/μm栅栏组织厚度Thickness of palisade tissue/μm海绵组织厚度Thickness of spongy tissue/μmCTR/%SR/%CK224.57c3.30bc72.33bc101.91c32.20ab45.38bT1245.21b2.42d82.22a118.81ab33.45a48.45aT2235.80b2.84cd65.28c115.24b27.72c48.84aT3257.13a4.55a82.72a113.00b31.99ab44.10bT4256.03a3.66b76.21ab125.41a29.68bc49.10aT5237.91b3.39b78.29ab103.82c32.90a43.62b
注:CTR表示叶片组织结构紧密度;SR表示叶片组织结构疏松度;同列不同小写字母表示不同处理P<0.05水平差异显著。以下图表同上。
Note:CTRmeans the leaf tissue structure tense ratio;SRmeans the leaf tissue structure spongy ratio; Different small letters in one column mean significant difference among treatments atP<0.05. The same as below.
2.2 叶片激素含量变化
由表3可知,与对照相比,5种处理均显著提高叶片中的ABA含量,其中T5含量最高,达到118.87 ng·g-1;除T5的IAA和GA含量显著低于对照外,其它处理均高于对照;T3和T4的ZR含量均显著高于对照,其它处理均显著低于对照,其中T5的ZR含量最低,仅7.3 ng·g-1。年灌水量相同(年灌水总量为5400 m3·hm-2)、灌水次数与下渗深度不同的T2与T3相比,T2的ABA、IAA和GA含量均显著高于T3,但T2的ZR含量显著低于对照,说明年灌水量相同,灌水次数和灌水深度对叶片中激素含量影响显著。
2.3 基因克隆及表达分析
2.3.1PbDREB基因全长cDNA克隆及序列分析 通过ORF Finder分析,PbDREB基因全长cDNA为1551 bp,其中包含5′非编码区10 bp,3′非编码区346 bp和开放阅读框1197 bp,编码398个氨基酸,起始密码子为ATG,终止密码子为TGA。

表3 不同灌溉处理对黄冠梨叶片ABA、IAA、GA和ZR含量(鲜重)的影响
通过Protparam分析其理化性质,推测PbDREB蛋白分子式C1858H2863N531O630S23,其相对分子量为43456.7,等电点4.8。其理论半衰期约为30 h,不稳定参数为44.58,蛋白质性质不稳定(40以下为稳定蛋白)。该蛋白中相对含量较多的氨基酸是甘氨酸Gly(11.3 %,45个)、丝氨酸Ser(10.1 %,40个)和谷氨酸Glu(8.5%,34个),预测该蛋白为亲水性蛋白。TMHMM-2.0分析PbDREB氨基酸序列没有跨膜结构域,没有信号肽。NetPhos2.0预测结果显示,PbDREB蛋白有18个Ser、7个Thr和3个Tyr,可能成为蛋白激酶的磷酸化位点。推断PbDREB蛋白可能定位于细胞核。
将PbDREB基因编码的氨基酸序列进行同源分析,结果发现,该氨基酸序列与很多植物DREB氨基酸序列具有较高的一致性(图2)。PbDREB编码的氨基酸序列与苹果MdDREB2-1编码的氨基酸序列一致性最高,为59.8%,其它依次是沙冬青AmDREB2C38.8%、草莓FvDREB2C-like36.4%、蓖麻RcDREB2C34.2%、黄瓜CsDREB2C-like33.8%、鹰嘴豆CaDREB2C-like31.3%、川桑MnDREB2A30.7%。

注:图中分支点的数字表示Bootstrap验证中基于1000次重复该节点可信度的百分比;标尺代表遗传距离。Note: The number on the branches indicate the reliability percent of Bootstrap values based on 1000 replications; the scale bar represents genetic distance.图2 不同植物DREB氨基酸序列的系统进化树分析Fig. 2 Phylogenetic tree of DREB amino acid sequences from different plants
2.3.2PbDREB在不同灌溉处理下的表达 由图3可知,PbDREB在6种处理的叶片中均有表达,除T1与对照差异不显著外,其它处理均显著高于对照,其中在T5中的表达量最高,达到2.75,是对照的2.89倍;当灌水下渗深度同为60 cm时(T1、T2、T4和T5),各处理叶片PbDREB基因表达量随年灌水量的增加而上升,且达到差异显著水平。年灌水量相同(年灌水总量为5400 m3·hm-2)、灌水次数与下渗深度不同的T2与T3相比,PbDREB在T2中的表达量略低于T3,但未达到差异显著水平,说明灌水量相同时,灌水次数和下渗深度对PbDREB基因表达量的影响不显著。

图3 不同灌溉处理下叶片中PbDREB的表达水平Fig. 3 Expression pattern of PbDREB in the leaves of six different irrigation treatments
3 讨 论
果树的抗旱能力是一种复合性状[17],指标的合理选择是抗旱性鉴定的关键。在叶片结构上,CTR和SR等指标与果树的抗旱性密切相关,这在葡萄[18]、核桃[19]、桃[11]、酸枣[20]、苹果[21]等果树上已得到证实。郑国琦等[22]研究发现,月灌水定额小于900 m3·hm-2时,随着灌水量的增加,叶片厚度、栅栏组织厚度和叶片结构紧密度显著增加。有关不同灌水处理对梨叶片解剖结构的影响尚未见研究报道。本研究结果发现,叶片厚度、栅栏组织厚度和叶片结构紧密度并没有随着灌水量的增加而增加,但灌水量最大的T5(灌水9次、下渗60 cm、年灌水总量为6750 m3·hm-2的覆膜滴灌)处理的叶片栅栏组织厚度/海绵组织的比值最大、CTR较大、SR最小。栅栏组织厚度/海绵组织的比值和CTR增大是植物对水分短缺的一种响应,该特征有助于CO2气体从气孔下室到光合作用场所的传导,又可抵消因气孔关闭和叶肉结构的变化所引起的CO2传导率的降低,从而提高植物对水分的利用率,表现出植物的抗旱适应性[23]。
内源激素是植物体生命活动的调节者,在逆境胁迫下,植物通过内源激素的变化如IAA、ZR和CTK浓度的减少和ABA浓度的升高来调节某些生理过程以达到适应逆境的效果,已经在许多果树等植物上得到证明[24]。本研究发现,T5(灌水9次、下渗60 cm、年灌水总量为6750 m3·hm-2的覆膜滴灌)处理的叶片中ABA增加的同时,IAA、ZR和CTK均减少,从而可以改变膜的特性,使气孔关闭,改变代谢氧的容量与膜相结合的酶的活性,促进降解过程并抑制合成过程[25]。但关于灌水量对植物内源激素的影响,目前报道较少。王永平[26]研究发现,玉米籽粒的 ABA、Z+ZR、GAs和IAA含量受水分影响明显,随着灌水量提高内源激素含量显著提高。但本研究发现,随着灌水量的增多,内源激素含量并没有规律性地增多,引起相关差异的原因,有待进一步试验研究。
周永斌[27]等研究表明,在不同的干旱胁迫条件下,转基因(一种DREB类基因)植株的抗旱能力明显高于受体对照。周伟[28]等研究发现,重组酵母菌株与表达VaDREB基因的转基因拟南芥株系均表现出明显增强的盐、干旱、低温和高温胁迫耐受性表型,基因表达分析结果表明,转基因拟南芥株系中抗逆相关功能基因的表达量出现了明显上调。施肖堃[29]证明对甘蔗转入EaDREB2B基因可以提高其耐旱性。综上结果证明VaDREB转录因子在植物抗逆调控过程中起着重要作用。本研究结果表明,DREB在6个不同灌溉处理的黄冠梨叶片中都有表达,并且与灌水次数和灌水量呈正相关,说明6种不同灌溉处理的黄冠梨都具有一定的抗逆性,其中在T5(灌水9次、下渗60cm、年灌水总量为6750 m3·hm-2的覆膜滴灌)处理的叶片表达量最高。然而,DREB类转录因子超表达在提高植株耐逆性的同时还会产生生长迟缓、植株矮化、产量下降等不良影响,这种负效应可以通过用胁迫诱导启动子或其它条件性启动子启动DREB基因的表达来加以克服[30]。这一步骤在今后有待于进一步的研究。
参考文献:
[1] 滕元文,柴明良,李秀根.梨属植物分类的历史回顾及新进展[J].果树学报,2004,21(3):252-257.
[2] 王迎涛,李 晓,李 勇,等.套袋黄冠梨果实花斑病发生与其组织结构变化的关系[J].西北植物学报,2011,31(06):1180-1187.
[3] 王杰.梨历史与产业发展研究[D].福州:福建农林大学, 2011.
[4] 王恩鹏.西北地区水资源现状与可持续利用对策的探讨[J].云南地理环境研究,2006,18(1): 92-96.
[5] 张治,田富强,钟瑞森,等.新疆膜下滴灌棉田生育期地温变化规律[J].农业工程学报,2011,27(1): 44-51.
[6] 李昭楠,李 唯,刘继亮,等.不同滴灌水量对干旱荒漠区酿酒葡萄光合及产量的影响[J].中国生态农业学报,2011,19(6):1324-1329.
[7] 李敏敏,安贵阳,郭 燕,等.不同灌溉方式对渭北果园土壤水分及水分利用的影响[J].干旱地区农业研究,2011,29(4):174-179.
[8] 赵昌杰,刘松忠,张 强.果树对干旱胁迫的响应研究进展[J].中国果树,2011,(4):60-62.
[9] 赵延霞,骆有庆,宗世祥,等.不同沙棘品种雌雄株叶片解剖结构及抗旱性比较[J].北京林业大学学报,2012,34(6):31-41.
[10] 孟庆杰,王光全,董绍锋,等.桃叶片组织结构与其抗旱性关系的研究[J].西北林学院学报,2005,20(1):65-67.
[11] 孟庆杰,王光全,董绍锋,等.桃叶片组织解剖结构特征与其抗旱性关系的研究[J].干旱地区农业研究,2004,22(3):123-126.
[12] 刘长海,周莎莎,邹养军,等.桃叶片组织解剖结构特征与其抗旱性关系的研究[J].干旱地区农业研究,2012,30(5):94-98.
[13] Sun J W, Peng X J, Fan W H, et al. Functional analysis of BpDREB2 gene involved in salt and drought response from a woody plant Broussonetia papyrifera [J]. Gene, 2014, 535(2):140-149.
[14] Se-jun O, Ik S S, Shic K Y, et al. Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth [J]. Plant Physiol, 2005, 138(1): 341-351.
[15] 樊卫国,李迎春.部分梨砧木的叶片组织结构与抗旱性的关系[J].果树学报,2007,25(1):17-21.
[16] 李宗霆,周 燮.植物激素及其免疫检测技术[M].南京:江苏科学技术出版社,1996.
[17] James A S, Sarah J G, William R G. Resistance to water stress of alnus maritima: intraspecific variation and comparisons to other alders [J]. Environ Exp Bot, 2005, 53(3): 281-298.
[18] 潘学军,张文娥,杨秀永,等.喀斯特山区野生葡萄实生苗叶片解剖结构与抗旱性的关系[J].贵州农业科学,2010,38(9):176-182.
[19] 李 军,卫发兴,陈风顺,等.从六个核桃无性系(种)叶的形态解剖比较其抗旱性[J].河南林业科技,1997,17(3): 9-11.
[20] 刘学师,任永信,任小林.酸枣叶片组织结构与抗旱性的研究[J].河南职业技术师范学院学报,2004,32(1):45-47.
[21] 马兰青,王丽雪,王有年,等.干旱对苹果叶片形态解剖结构的影响[J].北京农学院学报,1996,11(2):27-31.
[22] 郑国琦,张磊,郑国保,等.不同灌水量对干旱区枸杞叶片结构、光合生理参数和产量的影响[J].应用生态学报,2010,21(11):2806-2813.
[23] Vaia L, Artemios M B, Angelos P, et al. Effects of chilling stress on leaf morphology, anatomy, ultrastructure, gas exchange, and essential oils in the seasonally dimorphic plant Teucrium polium (Lamiaceae) [J]. Acta Physiol Plant, 2014, 36(8): 2271-2281.
[24] Yang D L, Yang Y N, He Z H. Roles of plant hormones and their interplay in rice immunity [J]. Mol plant, 2013, 6(3): 675-685.
[25] 张明生,谢 波,谈 锋.水分胁迫下甘薯内源激素的变化与品种抗旱性的关系[J].中国农业科学,2002,35(5):498-501.
[26] 王永平.灌溉和地膜覆盖对玉米籽粒灌浆的影响及其与激素的关系[D].杨凌:西北农林科技大学,2014.
[27] 周永斌,闵东红,徐兆师,等.不同水分胁迫下转W16小麦回交株系基因表达特性及其抗旱机制[J].农业生物技术学报,2012,20(11):1271-1281.
[28] 周伟,吴 宪,贾承国,等.山葡萄转录因子基因VaDREB的克隆及其抗逆功能分析[J].农业生物技术学报,2014,50(10):1563-1573.
[29] 施肖堃.转DREB基因甘蔗的抗旱性评价及其遗传稳定性分析[D].福州:福建农林大学, 2010.
[30] 阳文龙,刘敬梅,刘强,等.高羊茅DREB 类转录因子基因的分离及鉴定分析[J].核农学报,2006,20(3):187-192.
