福清龙江不同断面空心莲子草生理生态特征比较
2018-07-02黄相玲
黄相玲
(广西大学林学院,广西南宁 530004)
龙江是闽东独流入海的水系,流经福建省福清市城市规划区,是福清市最大的河流,自西向东流经东张、宏路、融城等地,于海口注入福清湾,其支流有大北溪、入城溪、虎溪等。龙江发源于莆田县境内,全长62 km,在福清境内全长35.51 km,流域面积583 km2,年平均径流量1.69亿m3。龙江是福清市经济发展极为重要的生态载体,但长期以来,水资源的过度开发利用、河道淤积、流域周边工农畜牧业以及沿江生活污水无序排放等生态破坏活动造成许多环境污染问题,从而给龙江流域植物的生长带来一定的影响。
空心莲子草(Alternantheraphiloxeroides)别称喜旱莲子草、水花生、空心苋水蕹菜、革命草,属苋科(Amaranthaceae)莲子草属(Alternanthera)多年生宿根草本植物,原产于南美洲,分布在巴拉圭、阿根廷南部巴拉那河流域和巴西南部沿海地带,20世纪30年代末日本侵华期间,曾被用作饲料在我国广泛引种,后逸为野生。空心莲子草具有极强的无性繁殖能力、抗逆能力与适应性,在水田、湿地和旱地中均可生长,在杂草的生存竞争中占有绝对优势。在入侵环境中空心莲子草缺乏种群依赖型自然天敌的控制,引入不久便大肆蔓延,造成严重的生态环境破坏和经济危害[1]。目前,我国除甘肃东南部、宁夏、陕西、山西、内蒙古南部以及辽宁南部尚未遭到空心莲子草入侵外,其他地区均有分布。空心莲子草已成为我国16种“危害最大的外来物种”之一,给我国的经济带来了巨大的损失[2-3]。
空心莲子草在福清龙江水域段是一种生长旺盛的外来入侵植物[4]。在龙江上有东张水库、天宝坡等水利工程,龙江在天宝坡以上为非感潮河段,在天宝坡以下为感潮河段,天宝坡下径流量很小。本研究以这3个不同断面空心莲子草为研究对象,对其生理生态进行分析,从而了解不同断面处空心莲子草生理生态的差异性,以期为龙江的环境污染治理提供基础资料。
1 材料与方法
1.1 试验地点概况
福清市龙江水域属于南亚热带海洋性季风气候,夏长暖湿,冬短温凉。年平均气温为19.7 ℃,最冷月(1月)平均气温为10.8 ℃,最热月(7月)平均气温为28.2 ℃,年平均降水量为 1 323.7 mm,一年中3—9月为多雨期,雨量约占全年的80%,10月至翌年2月为少雨期,土壤含水量约为60%,日照时数为1 984.3 h,日照百分比45%,年太阳辐射量为491.84 kJ,年平均风速为3.6 m/s。风速季节变化较明显,春夏风速小,秋冬风速大[4]。在龙江水域选择倪浦桥、观音埔过圳(天宝坡采样点)、瑞云桥3个典型的植物生长环境断面为试验点。
1.2 试验设计
试验于2016年10月初进行,以福清市龙江水域上游段倪浦桥、下游段观音埔、中游段瑞云桥3个不同断面空心莲子草为试验材料,每个断面处随机选取30株空心莲子草作为试验样品。用美国PP System公司生产的CIRAS2便携式光合作用测定仪测定空心莲子草叶片的光合指标。用英国Hansatech公司生产的Handy-PEA植物效率分析仪测定空心莲子草叶片荧光动力学指标。试验环境条件为野外自然环境条件,在气温、光照度相似的不同时间、不同断面采样、测量。用烘干称质量法测定空心莲子草植株地面上部分生物量。
1.3 生理生态指标的测定
1.3.1 生物量的测定 用试验钢卷尺测量空心莲子草整个植株的株高、叶长、叶宽等性状指标,叶面积采用便携式叶面积仪测量,将选取的30个样品的空心莲子草地面部分刈割采集后带回实验室置于烘箱于105 ℃、5 min杀青,然后于85 ℃、48 h烘干至恒质量,用精确至0.000 1 g的电子分析天平称其干质量。
1.3.2 叶片光合作用指标测定 用CIRAS2便携式光合作用测定仪测定空心莲子草叶片的光合指标,每株测定3张叶片,每个断面处测定30个个体样品,取平均值。测定参数包括:叶片净光合速率(Pn),单位为μmol/(m2·s);蒸腾速率(Tr),单位为mmol/(m2·s)、气孔导度(Gs),单位为 mmol/(m2·s);胞间CO2浓度(Ci),单位为 μmol/mol。测定时使用LED光源,控制光照度为 1 000 μmol/(m2·s),CO2浓度为自然浓度,温度为室外温度30 ℃,每个测试样稳定30 s后读数[5]。
1.3.3 叶片叶绿素荧光参数测定 在叶片自然生长角度不变的情况下,选取已测定过光合作用的空心莲子草叶片,用英国Hansatech公司生产的Handy-PEA植物效率分析仪测定空心莲子草叶片叶绿素荧光参数,主要有初始荧光(F0)、最大荧光(Fm)、最大可变荧光(Fv)、光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)、叶片性能指数(PIInst)、单位反应中心用于电子传递的光能(ET0/Rc)、单位反应中心传递到PSI的光能(RE0/Rc)和以吸收光能为基础的性能指数(PIabs)。测定前叶片均先暗适应30 min后方可开始测定[6]。
1.4 数据分析
运用Excel 2010软件对所有数据进行处理,利用SPSS 17.0数据统计分析软件对所有数据进行分析,采用单因素(One-ANOVA)方差分析及多重比较(采用LSD法)。
2 结果与分析
2.1 不同断面处空心莲子草生物量的变化
植物的叶片性状指标是植物生长情况的最直观表征。由表1可见,不同断面处空心莲子草的叶面积、株高、干质量存在显著差异,部分采样点间叶长、叶宽指标差异显著。叶长与叶宽在3个断面处表现出一致的规律:观音埔、倪浦桥2个断面采样点差异不明显,但与瑞云桥采样点差异显著。倪浦桥的空心莲子草的叶面积与瑞云桥断面处存在显著差异,但和观音埔断面采样点差异不显著;干质量指标中,观音埔断面处和瑞云桥、倪浦桥间均存在显著差异。经综合分析发现,瑞云桥断面处空心莲子草的叶片形态指标整体相对较高,生长占优势,表明瑞云桥断面空心莲子草可能受到的环境胁迫较小。

表1 不同断面处空心莲子生物量的变化Table1 The change of biomass of A. philoxeroides in cross-section habitats
注:同列数据后标有不同小写字母表示存在显著差异,“**”表示差异极显著(P<0.01),“*”表示差异显著(P<0.05)。下表同。
2.2 不同断面处空心莲子草光合指标的变化
植物的光合作用是植物积累能量、维持正常生长的主要途径之一[7-8]。由表2可知,在3个不同断面处空心莲子草的Pn、Gs、Ci指标均存在显著差异,而蒸腾速率指标存在显著差异。倪浦桥断面处空心莲子草的Pn显著大于瑞云桥断面处空心莲子草的Pn,但倪浦桥和观音埔差异不显著。倪浦桥、观音埔、瑞云桥3个断面处空心莲子草的蒸腾速率之间都存在显著差异。观音埔断面处空心莲子草的Gs显著大于倪浦桥和瑞云桥断面处的空心莲子草的Gs,但倪浦桥和瑞云桥差异不显著。倪浦桥和瑞云桥断面处空心莲子草的Ci差异显著但倪浦桥与观音埔断面处空心莲子草的Ci差异不显著。
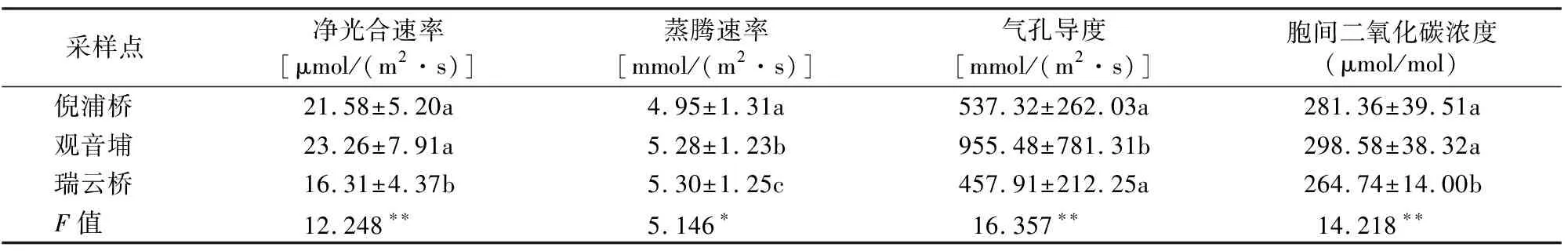
表2 不同断面处空心莲子草光合指标的变化Table2 The change of photosynthetic indexes of A. philoxeroides in cross-section habitats
2.3 不同断面处空心莲子草荧光参数的变化
由表3数据分析可知,经单因素方差分析,3个不同断面处空心莲子草F0、Fv/Fm值、Fm、Fv、F0/Fm值、ET0/Rc值、RE0/Rc值、PIabs、PIInst等参数在不同断面间存在显著差异。倪浦桥断面处空心莲子草的F0和RE0/Rc值显著大于观音埔与瑞云桥断面处,但观音埔和瑞云桥断面处这2个参数却无显著差异。Fv/Fm值、Fv、F0/Fm值、PIInst、PIabs这5个荧光参数指标在观音埔与倪浦桥2个断面之间存在显著差异,观音浦与瑞云桥在这5个荧光参数指标方面也差异显著,但倪浦桥断面与瑞云桥断面之间差异不显著。最大荧光Fm、ET0/Rc值在3个断面间均存在显著差异。
一般植物中Fv/Fm值在未受到胁迫的正常生长中是稳定的,常处于0.80~0.86之间[9]。而由表3可见,空心莲子草的Fv/Fm值在0.58~0.70之间,低于正常生长,说明空心莲子草受到某种环境因子胁迫。

表3 不同断面处空心莲子草荧光参数的变化Table 3 The change of fluorescence parameters of A. philoxeroides in cross-section habitats
3 讨论
空心莲子草原本主要生长在水生生境中,诸如湖泊、水湾等水体中,而随着入侵环境和人为活动的变化却扩散到水陆两栖生境,进而扩散到陆生生境[10]。空心莲子草PSⅡ最大光化学效率在未受到胁迫的植物中正常生长一般是稳定的,常处于0.80~0.86之间[11]。而研究结果表明,空心莲子草的Fv/Fm值在0.58~0.70之间,低于正常生长;初始荧光F0常用来度量色素吸收的能量中以热和荧光形式散失的部分,PSⅡ天线色素的热耗散常导致F0降低,而PSⅡ反应中心的破坏或可逆失活则能引起F0的增加。因此,可以根据F0的变化衡量PSⅡ的稳定性[12]。而研究发现3个不同断面处的空心莲子草F0增加幅度有差异,PSⅡ最大光化学效率值低于正常生长值,这进一步说明PSⅡ反应中心在不同断面处受到胁迫。而影响空心莲子草的PSⅡ最大光化学效率有诸多因素,如光照度、温度、水分、空心莲子草本身的酶活性等。
据研究,植物的叶绿素荧光特性因种类和生态环境的不同有很大的差异,如不同阔叶树种叶绿素荧光参数差异显著[13]。在遮阴与全光照下,濒危药用植物川贝母叶绿素荧光特性有明显不同[14]。本研究得出,空心莲子草在不同断面处之间的F0、Fm、F0/Fm值、Fv/Fm值、Fv、ET0/Rc值、RE0/Rc值、PIInst、PIabs9项叶绿素荧光参数表现出不同程度的差异。正常情况下,叶绿素吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散3种途径消化掉,这3种途径间存在此消彼长的关系,光合作用和热耗散的变化会引起荧光发射的相应变化,因此,荧光变化可以反映光合作用和热耗散的情况[15]。F0是固定荧光或初始荧光,它与叶片叶绿素浓度有关[16]。Fv代表可参与PSⅡ光化学反应的光能辐射部分,反映PSⅡ和原初电子受体Q的还原情况[17]。Fv/F0值表示PSⅡ的潜在活性,是PSⅡ原初光能转化效率,反映植物的潜在最大光合能力。本研究发现,在3个不同断面处,空心莲子草Fm、Fv/Fm值、F0/Fm值均有显著差异,所测的3个断面处空心莲子草种中,生物量干质量由大到小排序为倪浦桥>瑞云桥>观音埔,说明这3个断面处的空心莲子草的实际光合能力也是倪浦桥>瑞云桥>观音埔。
空心莲子草入侵成功的机制之一就是能够有效利用光能并使光合效率达到最大[18]。良好的光环境是杂草萌发和出苗的必备条件之一,但有试验结果表明,光对莲子草出苗的影响不显著[19]。光对营养生长时期的喜旱莲子草来说尤为重要。张彩云等的研究表明,25 ℃时喜旱莲子草的光饱和点、最大净光合速率和表观量子效率都显著高于莲子草[19]。高的光合速率是空心莲子草迅速生长和高光能利用率的基础,高的光饱和点有助于空心莲子草PSⅡ最大光化学效率的提高,但研究发现,空心莲子草的PSⅡ最大光化学效率低于其正常生长值,这说明空心莲子草光饱和点低于正常值。温度是导致这一结果的主要因素,本研究是于10月份在野外进行试验的,自然条件温度要低于春夏季节。温度降低直接影响空心莲子草的自身各种酶活性,间接影响其PSⅡ最大光化学效率。一般在强烈的太阳光照射后可激活空心莲子草自身内部过氧化氢酶[20],但在时间季节的选择下10月份的自然光照度通常要低于正常春夏季节光照度,研究表明影响空心莲子草的光合能力下降的关键因素是光照度和自然温度。
通过研究表明,空心莲子草的迅速生长繁殖不但与其生物学特性有关,而且与其周围环境有着密切的关系。龙江水域横跨区域较长,在不同河流段具有不同的河流性质和不同的人为活动造成的环境因子变化,给龙江水域水质、土壤、植物带来一定的负面的影响,空心莲子草是龙江水域的一种生命力较强、危害性较大的入侵植物,给龙江水域的生态系统及物种多样性带来不同程度的负面影响,需要对其进行防治以减少其危害。但目前具体是何种环境因素(如有机污染物、金属污染物或土壤基质营养性质等不同)引起空心莲子草生理生态特征在不同断面处的差异还不清楚,尚需进一步研究。综上所述,只有全面了解空心莲子草的生理生态特性变化才能制定出正确的防治措施。由此可见,空心莲子草的成功入侵是由多方面因素综合决定的,要综合考虑才能找出行之有效的方法去治理[21-23]。
参考文献:
[1]李 博,陈家宽. 生物入侵生态学:成就与挑战[J]. 世界科学技术研究与发展,2002,24(2):26-36.
[2]丁建清. 外来生物的入侵机制及其对生态安全的影响[J]. 中国农业科技导报,2002,4(4):16-20.
[3]徐承远,张文驹,卢宝荣,等. 生物入侵机制研究进展[J]. 生物多样性,2001,9(4):430-438.
[4]郑 丽,冯玉龙. 入侵植物的生理生态特性对碳积累的影响[J]. 生态学报,2005,25(6):430-1438.
[5]闫小玲,寿海洋,马金双. 中国外来入侵植物研究现状及存在的问题[J]. 植物分类与资源学报,2012,34(3):287-313.
[6]翁伯琦,林 嵩,王义祥. 空心莲子草在我国的适应性及入侵机制[J]. 生态学报,2006,26(7):2373-2381.
[7]耿宇鹏. 入侵种喜旱莲子草在异质生境中的适应对策研究[D]. 上海:复旦大学,2006.
[8]刘春花. 外来种喜旱莲子草的入侵生态学[D]. 武汉:武汉大学,2006.
[9]陈中义,江红英. 空心莲子草无性繁殖体的大小及数量对其建群效应的影响[J]. 长江大学学报(自然科学版)2009,6(1):72-75.
[10]李水清,胡小冬. 草甘膦在空心莲子草中的吸收与传导作用研究[J]. 长江大学学报(自然科学版),2008,5(4):1-4.
[11]孙方胜. 水花生化学防除技术[J]. 安徽农学通报,2005,11(2):58-64.
[12]谭 苹,张学俊,杨建明,等. 空心莲子草与氯硝柳胺复配灭螺作用的研究[J]. 中国媒介生物学及控制杂志,2008(5):439-441.
[13]谭 苹,肖少玉. 空心莲子草对斑马鱼的急性毒性研究[J]. 安徽农业科学,2010(8):4046-4048.
[14]谭 苹,张学俊,杨建明,等. 空心莲子草对钉螺酶组织化学的影响[J]. 中国寄生虫学与寄生虫病杂志,2009(1):11-16.
[15]周小平,王建国,薛利红,等. 浮床植物系统对富营养化水体中氮、磷净化特征的初步研究[J]. 应用生态学报,2005(11):195-199.
[16]刘弋潞,何宗健. 水生植物净化富营养化水质的机理探讨和研究进展[J]. 江西化工,2006(1):27-30.
[17]潘庆民,韩兴国,白永飞,等. 植物非结构性贮藏碳水化合物的生理生态学研究进展[J]. 植物学通报,2002,19(1):30-38.
[18]周国庆,彭友林,王 云. 洞庭湖区喜旱莲子草的分布、发生与危害[J]. 安徽农业科学,2007,35(22):6892-6894.
[19]张彩云,刘 卫,徐志防,等. 入侵种喜旱莲子草和莲子草的营养生长和光合作用对温度的响应[J]. 热带亚热带植物学报,2006,14(4):333-339.
[20]沈健英,黄渊智. 环境因子对空心莲子草出苗和生长的影响[J]. 上海农业学报,2006,22(1):34-38.
[21]周 强,周 浩,李特灵. 湘北地区草坪杂草种类、分布及其危害的评价研究[J]. 草业科学,2004,21(3):59-62.
[22]周小刚,杨兴有,阳苇丽,等. 达州市烟田杂草种类及群落数量分析[J]. 杂草学报,2016,34(2):12-16.
[23]何维明. 水分因素对沙地柏实生苗水分和生长特征的影响[J]. 植物生态学报,2001,25(1):11-16.
